Article Text

Statistics from Altmetric.com
Editor—Silver-Russell syndrome (SRS) is a condition characterised by intrauterine and postnatal growth retardation with relative sparing of cranial growth, triangular facies, fifth finger clinodactyly, and facial, limb, or truncal asymmetry.1 2 The molecular basis of SRS remains elusive and seems likely to be heterogeneous. However, maternal uniparental disomy of chromosome 7 (mUPD7) has been found in approximately 10% of SRS patients, suggesting that at least one gene on chromosome 7 is imprinted and involved in the pathogenesis of this condition.3 4 Interest has surrounded the human chromosomal region 7p12-13, which is homologous to mouse proximal chromosome 11, since mUPD for this region in mice leads to prenatal growth failure.5 Within this region lie the genes for insulin-like growth factor binding proteins 1 and 3 (IGFBP1 andIGFBP3). Both are involved in regulation of fetal growth via the insulin-like growth factor axis.6 IGFBP1 and IGFBP3are therefore obvious candidates for a role in the SRS phenotype associated with mUPD7.3 4
In order to determine whether IGFBP1 and/orIGFBP3 are likely to be implicated in SRS, we have investigated their imprinting status in normal fetal tissues collected from termination of pregnancies. Samples were obtained from Queen Charlotte's and Chelsea Hospital with the approval of the Research Ethics Committee of the Royal Postgraduate Medical School (96/4955) and from the MRC Tissue Bank at Hammersmith Hospital (L Wong). Genomic DNA and total RNA was extracted from these tissues and RT-PCR performed as previously described.7 The primers used in this study are listed in table 1. PCR was carried out for 35 cycles for all reactions. Products were sequenced on an ABI PRISM 377 automated DNA sequencer using the dRhodamine Ampli-Taq dye-terminator cycle sequencing kit, according to the manufacturer's instructions.
Oligonucleotide primers used to investigate the imprinting status of IGFBP1 and 3
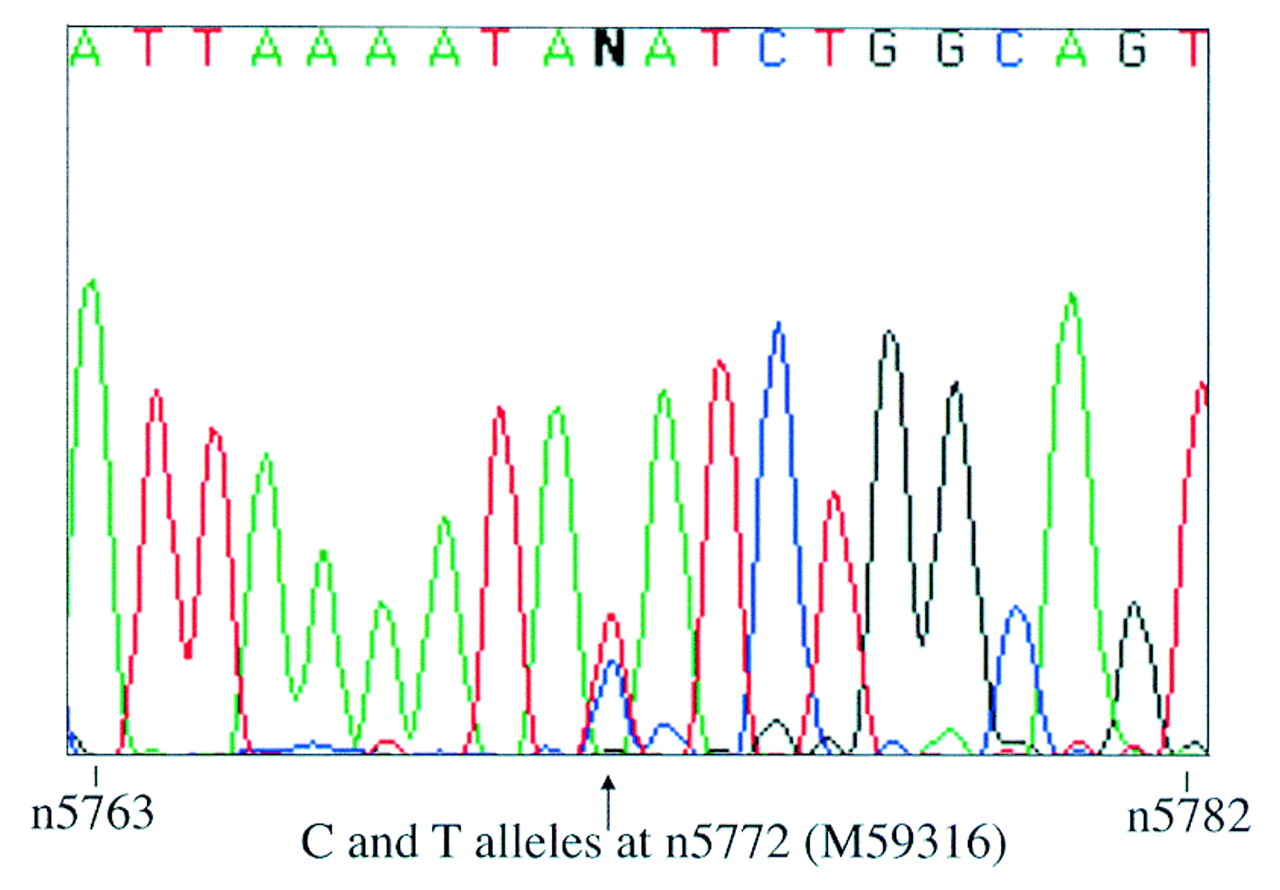
Since fetal IGFBP1 expression is highly tissue specific,8 its imprinting status was only investigated in fetal liver. Primers IGFBP1-BF and -ER were designed to span an expressed polymorphism at nucleotide 5772 within exon 4 (Genbank accession: M59316).9 These primers were used to screen genomic DNA from 16 first and second trimester fetuses. Five were identified as being heterozygous for the polymorphism and thus informative. Their gestational ages ranged from 5 to 17 weeks. Primers IGFBP1-EF and -ER, which span introns 2 and 3, were used to amplify liver cDNA derived from these informative fetuses. Control samples prepared from liver RNA without the addition of reverse transcriptase (RT) were also amplified. No genomic contamination was observed in any of the samples. RT-PCR products were sequenced using the reverse primer IGFBP1-ER and biallelic expression was seen in all five cases (fig1).
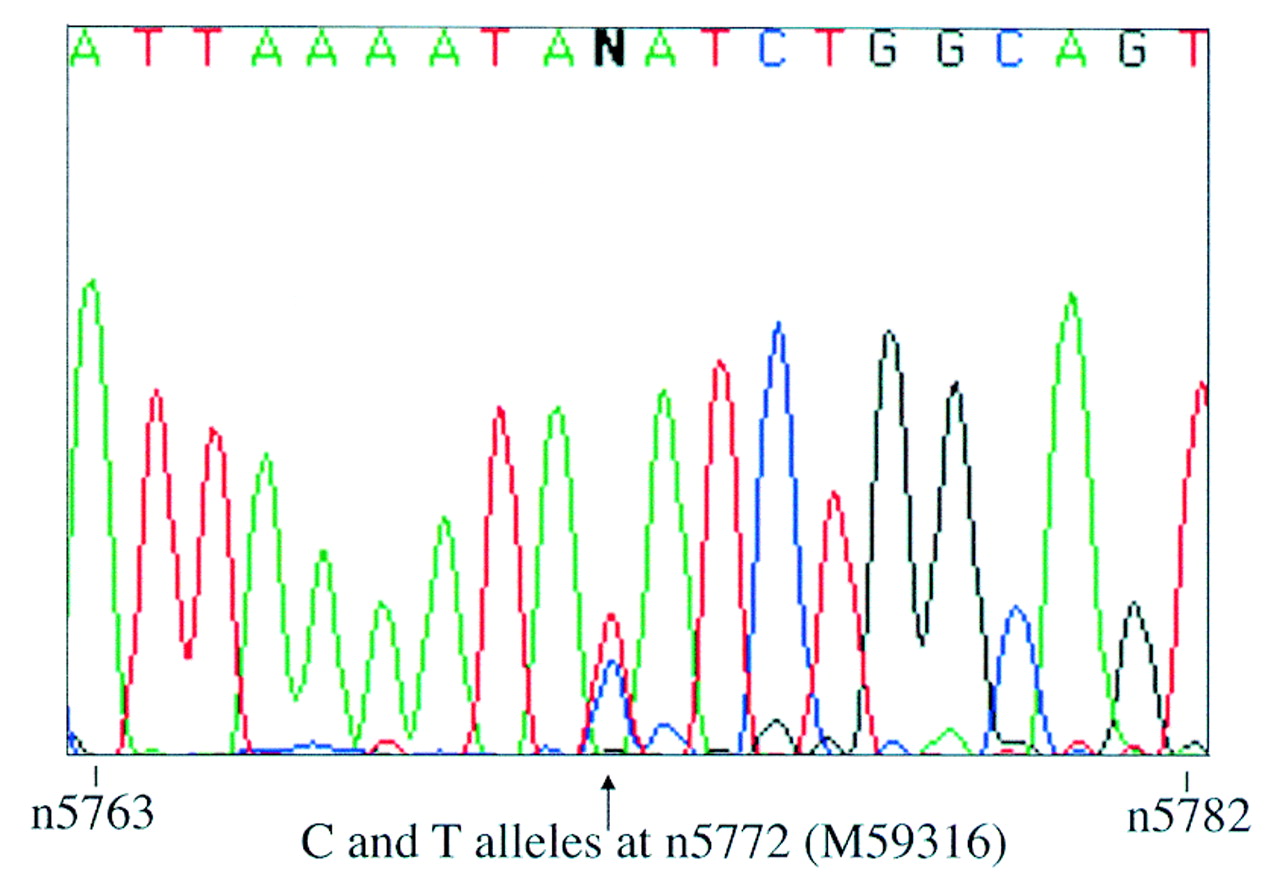
Biallelic expression of IGFBP1 in liver from one 12 week heterozygous fetus. Sequence is shown in the reverse complement direction and the polymorphic site is indicated by an arrow.
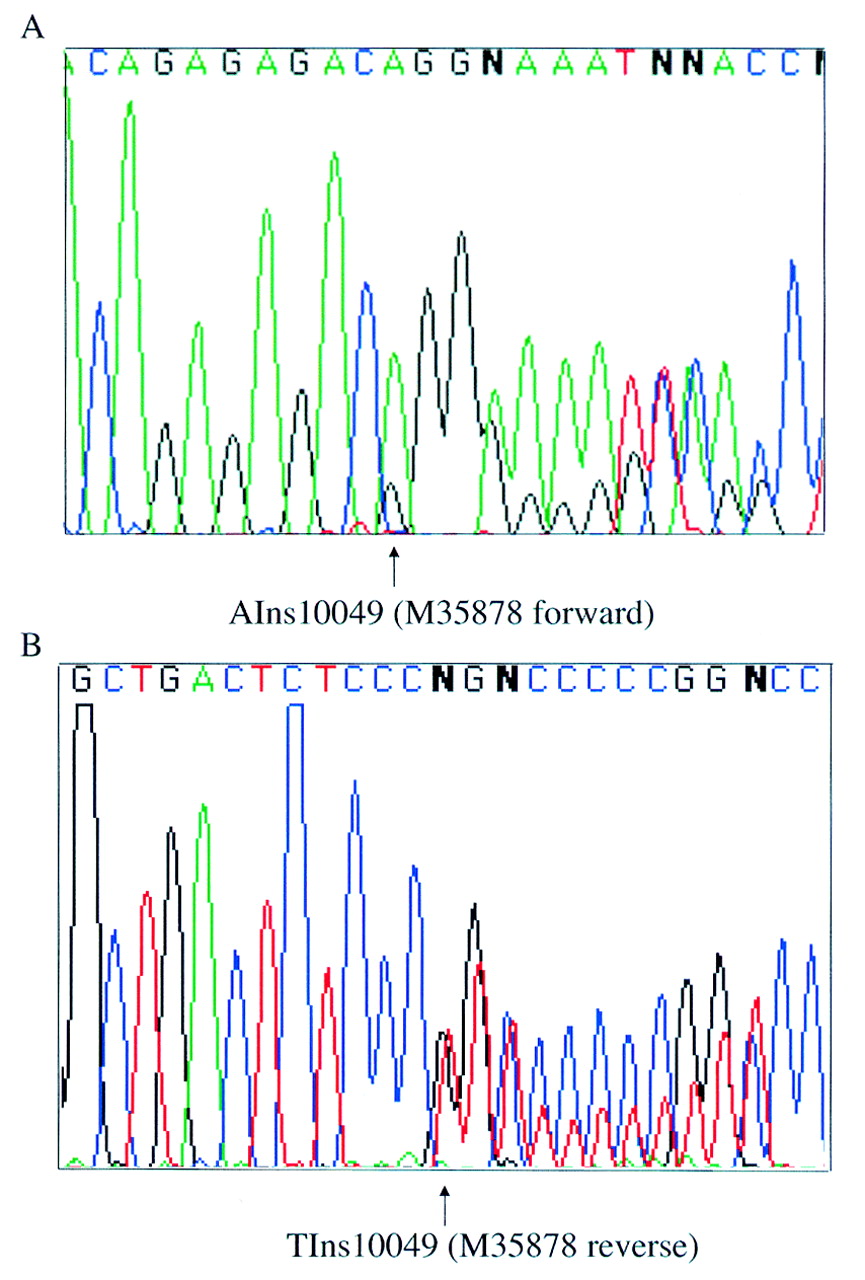
An expressed polymorphism, consisting of an adenine insertion at nucleotide position 10049 (Genbank accession: M35878) within the 3′ untranslated region of IGFBP3 (Dr I Morison, personal communication), was used to study the parental origin of expression in this gene. Primers IGFBP3-EF and -ER, from within exon 5, span this polymorphism and were used to screen fetal genomic DNA. The heterozygosity for this sequence change was 21%, with the adenine insertion being the most frequent allele in a UK based population. A total of 21 samples were screened and three heterozygous fetuses of 8-10 weeks' gestation were identified. Paired maternal DNA was homozygous in each case. Since IGFBP3 is widely expressed,8 parental allele expression was examined in brain, placenta, limb, and skin from all three fetuses. cDNA was amplified by RT-PCR using primers IGFBP3-GF and -ER which span intron 4. RT-PCR products were sequenced in both forward and reverse directions. Absence of genomic contamination was confirmed by using RT negative control samples and by sequencing RT-PCR products across the boundary of intron 4. Both parental alleles were detected in every tissue examined for all three fetuses (fig 2).
{kind=link}
{kind=link}
Biallelic expression of IGFBP3 in brain from one 10 week heterozygous fetus. Sequence is shown in both the forward (A) and reverse (B) directions. The polymorphic site is indicated by arrows.
The expression of IGFBP3 was also investigated in fibroblast and transformed lymphoblast cells from SRS patients. RNA was obtained from two normal subjects, two SRS patients with mUPD7, and four SRS patients without mUPD7.IGFBP3 mRNA expression was studied by RT-PCR using primers IGFBP1-F and -R. Products of the expected size (301 bp) were seen in UPD and non-UPD patients, as well as in the normal controls (data not shown). Similar results were obtained using transformed lymphoblast cells. Since IGFBP1expression was not detectable in either transformed lymphoblasts, lymphocytes, or fibroblasts, mRNA in mUPD7 patients could not be similarly investigated. However, IGFBP1 and IGFBP3 were both present in the serum of the two mUPD7 patients at normal levels.
These results show that in fetal tissues bothIGFBP1 and IGFBP3are biallelically expressed. Expression ofIGFBP3 in fibroblasts and lymphoblasts from patients with mUPD7 also rules out paternal specific expression of this gene in these cells in childhood. Although the expression ofIGFBP1 could not be directly investigated in SRS patient cell lines, because of its tissue specificity, serum levels in mUPD7 patients were normal. We therefore found no evidence for imprinting of IGFBP1 orIGFBP3, making their involvement in SRS unlikely.
Tissue and developmental stage specific imprinting is being reported in an increasing number of genes including KCN8(KvLQT1), IGF2,CDKN1C(p57Kip2 ),UBE3A, andMEST. 10 These make absolute exclusion of a gene as a candidate, on the basis of lack of observable monoallelic expression, difficult. Allelic expression ofIGFBP1 and IGFBP3was investigated during the fetal period and in tissues that were predicted to be biologically relevant. However, the possibility of imprinting of either gene in a particular cell type or restricted window of time could not be ruled out. Although fetuses of up to 18 weeks' gestation were screened for IGFBP3heterozygosity, the latest stage at which an informative fetus was identified was 10 weeks gestation. Demonstration ofIGFBP3 biallelic expression was therefore limited to the first trimester.
Quantitative analysis was attempted using SSCP analysis but neither polymorphism could be detected by this method. As the relative level of transcripts from the two alleles was not determined, preferential expression from one parental allele in either gene was not excluded.
Contamination by maternal tissue could potentially obscure an imprinting effect in fetal samples. For tissues used to studyIGFBP3 expression, paired maternal blood samples were available. All three maternal samples from heterozygous fetuses were found to be homozygous for the polymorphism studied. Maternal specific expression could therefore not have been missed. Although it is possible that maternal contamination of fetal tissue is obscuring paternal specific expression, the demonstration of biallelic expression in all samples studied makes this unlikely. Paired maternal tissue was not available for fetal liver tissues used in the study ofIGFBP1. However, the consistent observation of both alleles in five different samples makes it unlikely that monoallelic expression is being masked by maternal contamination.
Several other candidates for SRS have already been excluded. The involvement of IGF1R in SRS is unlikely since no hemizygosity or mutations were found for this gene in a cohort of 33 SRS patients.11 Riesewijk et al 12 have also failed to find evidence for a role of MEST, an imprinted gene located at 7q32. Two other candidates lie close to IGFBP1 andIGFBP3 in the region 7p12-13.EGFR is biallelically expressed and therefore unlikely to play a role in SRS.6 GRB10 is homologous to mouseMeg1/Grb10, a gene recently identified in a systematic screen for maternally expressed imprinted genes by subtraction hybridisation.13 The imprinting status ofGRB10 in man remains to be determined and its role in SRS is currently under investigation. Since imprinted genes tend to be located in clusters, it is likely that other imprinted genes lie within the regions surrounding MEST and probably, also, GRB10. These regions remain of interest in the search for candidate genes for SRS.
Acknowledgments
We thank the medical staff at Queen Charlotte's Hospital for help collecting samples and Sue Blunt from the cytogenetics laboratory for culturing fibroblast cells. This work was supported by funding from Action Research (EW), The Dunhill Medical Trust (SA), and Children Nationwide and Pharmacia and Upjohn (MH). Information regarding theIGFBP3 polymorphism was obtained from Dr I Morison, Department of Biochemistry, University of Otago, New Zealand.