Article Text

Abstract
Background TUBB8 is a primate-specific β-tubulin isotype whose expression is confined to oocytes and the early embryo. We previously found that mutations in TUBB8 caused oocyte maturation arrest. The objective was to describe newly discovered mutations in TUBB8 and to characterise the accompanying spectrum of phenotypes and modes of inheritance.
Methods and results Patients with oocyte maturation arrest were sequenced with respect to TUBB8. We investigated the effects of identified mutations in vitro, in cultured cells and in mouse oocytes. Seven heterozygous missense and two homozygous mutations were identified. These mutations cause a range of folding defects in vitro, different degrees of microtubule disruption upon expression in cultured cells and interfere to varying extents in the proper assembly of the meiotic spindle in mouse oocytes. Several of the newly discovered TUBB8 mutations result in phenotypic variability. For example, oocytes harbouring any of three missense mutations (I210V, T238M and N348S) could extrude the first polar body. Moreover, they could be fertilised, although the ensuing embryos became developmentally arrested. Surprisingly, oocytes from patients harbouring homozygous TUBB8 mutations that in either case preclude the expression of a functional TUBB8 polypeptide nonetheless contained identifiable spindles.
Conclusions Our data substantially expand the range of dysfunctional oocyte phenotypes incurred by mutation in TUBB8, underscore the independent nature of human oocyte meiosis and differentiation, extend the class of genetic diseases known as the tubulinopathies and provide new criteria for the qualitative evaluation of meiosis II (MII) oocytes for in vitro fertilization (IVF).
- TUBB8
- mutations
- oocyte maturation arrest
- female infertility
Statistics from Altmetric.com
Introduction
An essential feature of successful mammalian reproduction is the fusion of a sperm with a metaphase II oocyte.1 ,2 Oocytes are first arrested at prophase I and resume meiosis in response to luteinising hormone.3 ,4 Following spindle assembly and extrusion of the first polar body, oocytes are again arrested at metaphase II until fertilisation.5 ,6 This complex process is regulated by several signalling pathways.7–14 A number of mouse genes have been identified with mutations that cause an oocyte maturation arrest phenotype.15–19 However, in humans, only a few cases of primary infertility caused by oocyte maturation arrest have been reported,20–25 and the genetic aetiology of human oocyte maturation arrest is still largely unknown.
Although the mammalian oocyte meiotic spindle differs from the mitotic spindle in somatic cells in several respects,26 ,27 microtubules are the principal structural components in each case. These are dynamic filamentous polarised polymers assembled from α/β tubulin heterodimers; the latter consist of one α-tubulin and one β-tubulin polypeptide in tight association with one another. The productive folding of α-tubulin and β-tubulin and their integration into heterodimers cannot occur spontaneously. Rather, these processes require the interaction of newly synthesised polypeptides with several molecular chaperones. These include Prefoldin, which binds to and stabilises nascent α-tubulin and β-tubulin polypeptides;28 ,29 the cytosolic chaperonin, CCT,30 which provides a sequestered environment in which non-native tubulins can partition to a quasi-native conformation in the absence of off-pathway interactions that would otherwise lead to aggregation;31 and five tubulin-specific chaperones termed TBCA-E that function in concert as a GTP-dependent heterodimer assembly nanomachine.32
The α-tubulin and β-tubulin are each encoded by a multigene family containing seven (α) or nine (β) members. Each encoded protein is termed an isotype; these are highly homologous in amino acid sequence and differ from other family members by a relatively small number of conservative amino acid substitutions throughout the polypeptide as well as a more distinctively different acidic C-terminal tail.33 The amino acid sequence of each isotype is rigidly conserved across species boundaries, and the expression patterns of different α-tubulin and β-tubulin isotypes vary both as a function of development and in a tissue-specific manner. Not surprisingly, therefore, there is mounting evidence that the isotype composition of microtubules can contribute to their functional behaviour.33–36 The essential biological contribution of various α-tubulin and β-tubulin isotypes is reflected in the recent discovery of a large number of naturally occurring mutations in the genes that encode them; these mutations cause a range of mostly devastating developmental disorders collectively termed tubulinopathies.37–39
Among the isotypes encoding β-tubulin, TUBB8 is unusual in that it exists only in primate species, where its biological function was unknown. Recently, we identified seven missense mutations in TUBB8 in patients with oocyte maturation arrest.40 We found that TUBB8 is uniquely expressed in oocytes and the early embryo, where it is the preponderant β-tubulin isotype. Based on an assessment of their effects upon expression in HeLa cells, yeast cells, mouse and human oocytes, we concluded that these mutations exert their effects via dominant negative effects on microtubule behaviour. These findings uncovered an essential role for TUBB8 in human oocyte maturation and female fertility.
Here we report the identification of nine novel TUBB8 mutations in patients with oocyte maturation arrest. Among seven missense mutations, three (S176L, V255M and R262W) are de novo (of which one—S176L—is recurrent), one (T238M) is inherited and three (I210V, T285P and N348S) are of an unknown inheritance pattern. We also identify two homozygous patients with either an internal deletion of seven amino acids (E27_A33del) or a deletion leading to a frame shift (T143Dfs*12) in TUBB8, in either case precluding the expression of a functional TUBB8 polypeptide. Surprisingly, oocytes from patients with homozygous mutations each have a visible spindle, implicating spindle microtubule assembly from pre-existing non-TUBB8 isotypes expressed at an earlier developmental stage. Equally surprisingly, oocytes from patients with certain TUBB8 missense mutations (I210V, T238M and N348S) are capable of extruding the first polar body in the absence of a properly assembled spindle; these oocytes can be fertilised and undergo cleavage to generate embryos that fail to develop beyond the four-cell or eight-cell stage. These key findings dramatically extend the dysfunctional phenotypes of oocytes caused by mutation in TUBB8 and demonstrate that in humans oocyte differentiation occurs independently of meiosis.
Materials and methods
Human subjects and permission for animal experimentation
Nine families with oocyte maturation arrest were referred from the reproductive medicine centre at Ninth Hospital affiliated with Shanghai Jiao Tong University and Shanghai Ji Ai Genetics. All studies on human subjects were approved by the Fudan University Institutional Medical Review Board. A 32-year-old patient (body mass index index 23.2) had a single attempt at intracytoplasmic sperm injection (ICSI) because of problems relating to her husband's sperm quality. Four immature oocytes and seven MII oocytes were retrieved. Control MII oocytes for immunostaining were from four immature oocytes undergoing maturation in vitro, while the seven MII oocytes were used either for morphological evaluation by light or polarisation microscopy, or for fertilisation and embryonic development. MI oocytes were retrieved from patients diagnosed with oocyte MI arrest and were cultivated for maturation in vitro for about 20–24 hours, followed by fixation for immunostaining. All oocytes from normal controls and patients were obtained with written informed consent signed by donor couples. The study was also approved by the Reproductive Study Ethics Committee of the Ninth Hospital affiliated with Shanghai Jiao Tong University. All protocols for the experimental use and euthanasia of mice were reviewed and approved by the Medical College of Fudan University and were in accordance with the Association for Assessment and Accreditation of Laboratory Animal Care Guidelines.
Sequencing analysis of TUBB8
Patient genomic DNA samples and those from their family members and controls were extracted from peripheral blood using standard methods. All exons and splicing sites of TUBB8 were amplified. Amplified fragments were directly sequenced using an ABI 3100 DNA analyzer (Applied Biosystems, Foster City, California, USA).
Evaluation of phenotypes of oocytes
Oocytes obtained from donors undergoing routine clinical ICSI were viewed by phase contrast and polarisation microscopy with an OLYMPUS IX71 inverted microscope system. A method of oocyte immunostaining was adapted from that previously described.40 Briefly, oocytes were first fixed in 1.5% paraformaldehyde containing 0.1% bovine serum albumin (BSA). Meiotic oocyte spindles were stained with an anti-ß-tubulin antibody (1:400, F2043, Sigma-Aldrich) or an anti-α-tubulin antibody (1:500, T9026, Sigma-Aldrich). In microinjection experiments, mouse oocytes were tested using the above anti-ß-tubulin antibody and an anti-FLAG antibody (1:500, A9594, Sigma-Aldrich). Hoechst 33342 (1:600, BD) was used to label DNA. Oocytes were mounted on glass slides and examined using a Leica TCS SP8 confocal laser-scanning microscope platform.
Molecular modelling
A high-resolution cryo-EM structure of the microtubule41 (PDB ID: 3JAS) was used to map the TUBB8 mutations described in this study onto the atomic model of the α/ß-tubulin heterodimer.
Expression of wild-type and mutant forms of TUBB8 in cultured cells
Constructs engineered for the expression of C-terminally FLAG-tagged TUBB8 (wild-type and mutant) were expressed by transfection into HeLa cells and their microtubule phenotypes examined by immunofluorescence as described previously.40
Folding kinetics of wild-type and mutant forms of TUBB8
Constructs encoding full-length wild-type or mutant forms of TUBB8 in plasmid vectors containing a T7 promoter were used to drive expression in a rabbit reticulocyte lysate cell free system (coupled transcription and translation (TNT), Promega) supplemented with 35S-methionine. The kinetic analysis was done and reaction products analysed either by sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE) or on non-denaturing polyacrylamide gels as described previously.40 Reactions designed to ‘chase’ folding intermediates into heterodimers by providing a source of unlabelled α-tubulin were done by adding unlabelled native tubulin heterodimers to coupled TNT reactions after 90 min, and the continuing the incubation for a further 60 min as described previously.32
Mouse oocyte collection, meiotic maturation and culture
Germinal vesicle (GV)-stage oocytes were isolated from the ovaries of 4–6-week-old female ICR mice cultured in M16 medium and collected for immunostaining as described previously.40
Expression constructs, generation of cRNAs and microinjection into mouse oocytes
Site-directed mutagenesis, transcription of full-length TUBB8 cDNAs contained in a vector that included a cytomegalovirus (CMV) promoter and a C-terminal FLAG tag, and microinjection of wild-type and mutant cRNAs into mouse GV oocytes was done as described previously.40
Results
Mutations in TUBB8 result in a variety of oocyte phenotypes
We recruited 10 patients from 9 independent and hitherto uncharacterised families that included one or more female members with persistent infertility. This resulted in the identification of nine mutations in TUBB8, of which eight are novel (figure 1). S176L is a recurrent de novo mutation whose occurrence we reported previously.40 T238M is an inherited heterozygous missense mutation, while V255M and R262W are de novo mutations. Patients with homozygous mutations have either an internal deletion of seven amino acids (E27_A33del) or a deletion (T143Dfs*12), resulting in a truncated polypeptide of 154 amino acids. Because of a lack of parental information regarding their fathers, I210V, T285P and N348S are mutations with an unknown inheritance pattern. The location of all these mutations as well as those previously identified within TUBB840 is shown in online supplementary figure S1A; all residues implicated are strictly evolutionarily conserved among primate species (see online supplementary figure S1B).
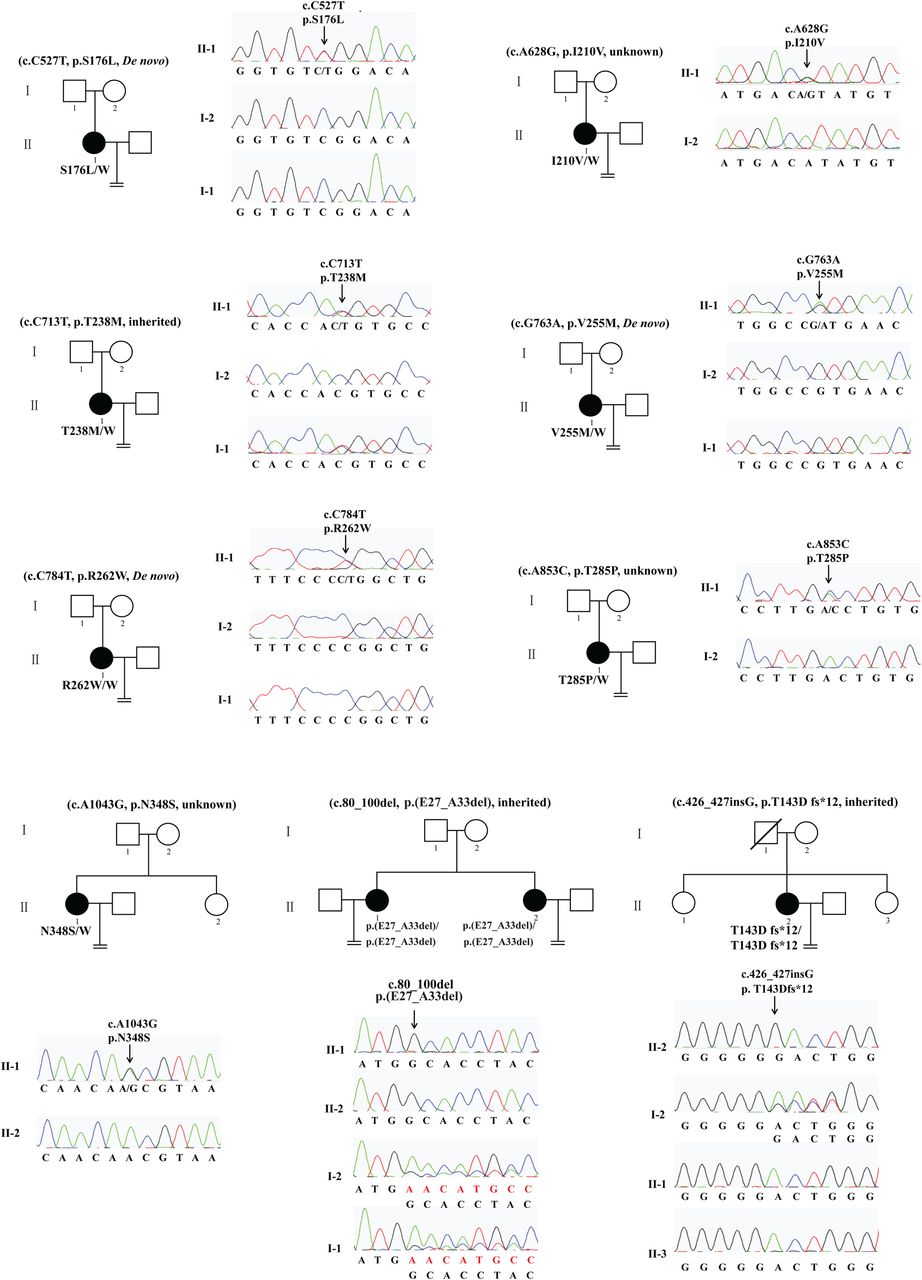
Pedigree and TUBB8 mutations leading to oocyte meiotic arrest. Pedigrees of nine families with mutations in TUBB8. The S176L, V255M and R262W mutations are de novo. The T238M mutation is inherited from her father, while the E27_A33del mutation and the T143Dfs*12 mutation are both homozygous, in each case inherited from heterozygous parents. The I210V, T285P and N348S mutations have unknown inheritance patterns. Sanger sequencing chromatograms are shown defining the nature and location of the mutations in each family. Squares denote male family members and circles female family members. Black circles represent affected individuals. Slash indicates deceased individual. An equal sign denotes infertility.
Supplemental material
All patients had primary infertility over the course of several years; their spouses had normal sperm counts, sperm morphology and motility. Each patient underwent 2–7 failed IVF/ICSI attempts (table 1). Specific information on the stage of oocytes retrieved is summarised in online supplementary table S1. We found several significant phenotypic differences among the patients' oocytes. Patients S176L, V255M, R262W and T285P had no morphologically identifiable MII oocytes (see online supplementary figure S2B, E, F and table S1). However, in patients I210V, T238M, E27_A33del and T143Dfs*12, there were some morphologically identifiable MII oocytes with an extruding first polar body (see online supplementary figure S2C, D, G, H). The relative proportion of morphological MII oocytes differed among these patients. The I210V patient had the highest proportion of MII oocytes (4/15), and in her latest attempt at ICSI, four out of five oocytes retrieved were morphologically MII with an extruding first polar body (figure 2C). In the latest ICSI attempt with the T238M patient, two out of nine oocytes retrieved were morphologically MII, with an extruding first polar body and no visible spindle (figure 2D). Upon immunostaining, oocytes of patients with heterozygous mutations had no detectable spindle (S176L, I210V, V255M and T285P), a conspicuously disorganised spindle (T238M), or a spindle that gave only a barely detectable immunofluorescence signal with an anti-β-tubulin antibody (N348S) (figure 2I).
Clinical characteristics of oocyte maturation arrest patients from the nine families

Oocyte phenotypes from patients with maturation arrest. A normal oocyte (A) and oocytes from patients in the families indicated (B-H) were separated from granulosa cells and examined by light and polarisation microscopy. Note that a normal MII oocyte has a first polar body (black arrow in A) and that normal MI and MII oocytes have visible spindles (white arrows in A). Patient oocytes S176L (B), V255M (E) and T285P (F) are at MI and none have a first polar body or a visible spindle. Some oocytes in the patient I210V (C) and T238M (D) have a first polar body (black arrow in C and D), but none of the oocytes from this patient have a visible spindle. All oocytes from patients E27_A33 del and T143Dfs*12 have a visible spindle (G and H), and an oocyte from the patient T143Dfs*12 has a first polar body (H). (I) Oocytes from control and various patients (n=1 for S176L; n=1 for T210V; n=2 for T238M; n=1 for V255M; n=2 for T285P; n=3 for N348S; n=2 for E27_A33del) were immunolabelled with antibodies against β-tubulin (to visualise the spindle, shown in green) and counterstained with Hoechst 33342 (shown in blue) to visualise DNA. No oocytes from the T143DFs*12 patient were available for this analysis and the spindle in the oocyte from the N348S patient is only very weakly visible compared with the wild-type control.
We previously established that TUBB8 is by a large margin the most abundant β-tubulin isotype expressed in human oocytes.40 In spite of the fact that the two homozygous mutations (p.T143Dfs*12 and p.(E27_A33del)) are incapable of generating a functional β-tubulin polypeptide (see below), oocytes harbouring these two mutations had a visible spindle by polarisation microscopy, although most of them lacked a first polar body (figure 2G,H). To see whether the absence of any functional TUBB8 in p.T143Dfs*12 oocytes might be compensated by an increase in expression of non-TUBB8 β-tubulin isotypes, we measured the levels of non-TUBB8 mRNAs in a single p.T143Dfs*12 oocyte, but found no such increase (data not shown). In addition, compared with a normal oocyte, immunofluorescence analysis of a p.(E27_A33del) oocyte showed the spindle to be less compact, with fewer polar microtubules (and possibly a lack of kinetochore microtubules) compared with a normal oocyte (figure 2I). We conclude that different TUBB8 mutations contribute to an extensive variety of oocyte phenotypes with respect to extrusion of the first polar body and spindle morphology, and that TUBB8 expression is essential for proper primate spindle morphology and function.
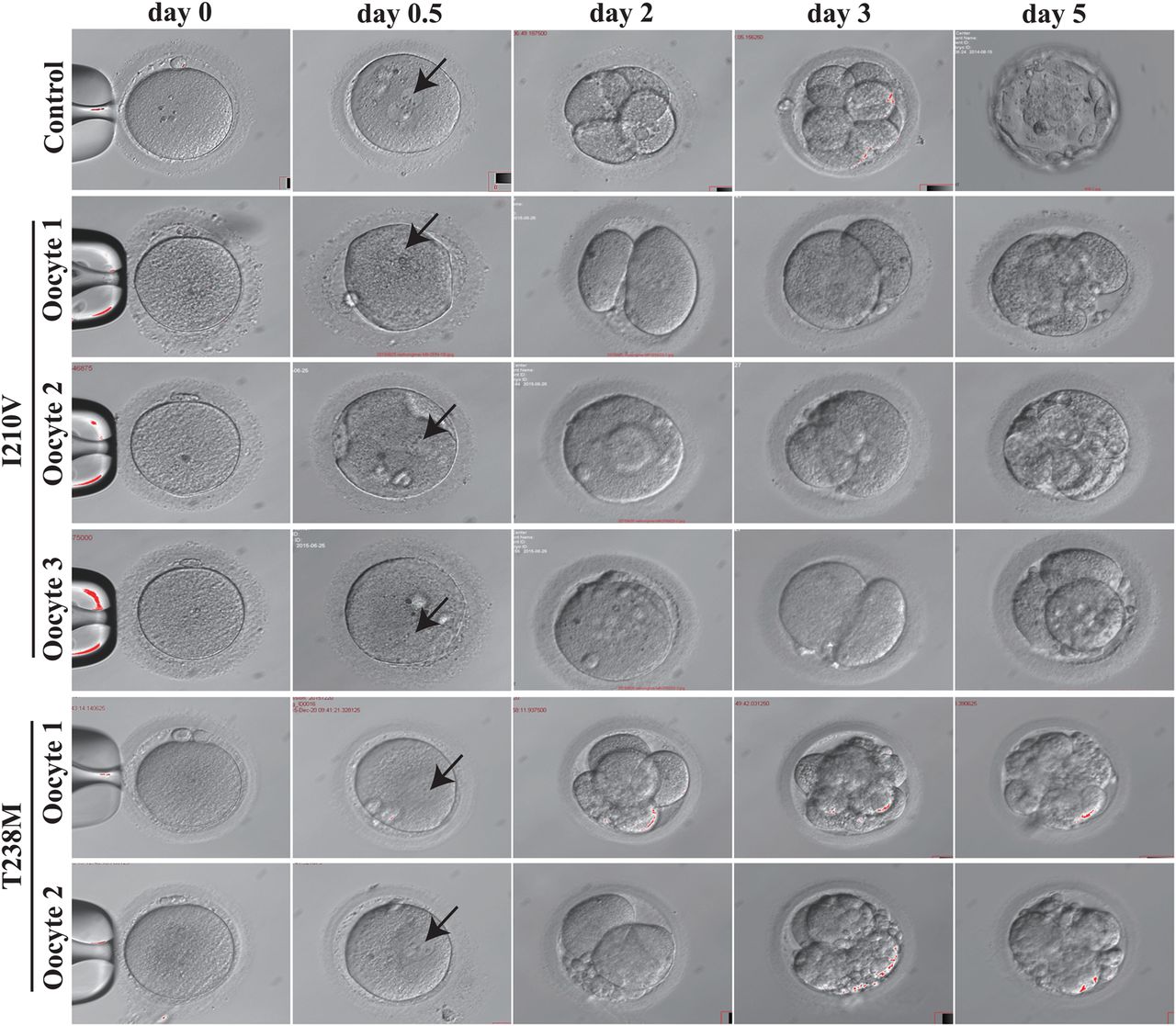
While many patient MII oocytes had no visible spindle by polarisation microscopy, we were surprised to discover that in her latest IVF attempt three out of four MII oocytes from the I210V patient could nonetheless be fertilised and underwent cleavage. However, all embryos became arrested at the two-cell stage on day 3, and by day 5 there were no viable embryos (figure 3). In the case of MII oocytes from the T238M patient, although two were abnormally fertilised in the sense that they had a 1PN pronucleus, each nonetheless underwent cleavage and formed either a class III eight-cell embryo or a class III four-cell embryo on day 3. Thereafter, in both cases, the embryos became arrested on day 5 (figure 3). The patient with the N348S mutation had a similar phenotype: eight oocytes became fertilised, but all embryos were non-viable by day 3 (data not shown). These observations further extend the range of dysfunctional oocyte phenotypes incurred by mutation in TUBB8 (summarised in table 2).
Spindle status, polar body extrusion, fertilisation and ensuing embryonic development in wild-type and patient oocytes harbouring mutations in TUBB8

Morphology of wild-type, I120V and T238M MII oocytes and early embryos. The morphologies of control MII oocytes (n=1), MII oocytes (n=3 for I210V; n=2 for T238M) from patients harbouring the indicated mutations in TUBB8, fertilised oocytes (on day 0.5), and embryos on days 2, 3 and 5 were examined by phase contrast light microscopy. Black arrows indicate the pronucleus.
Structural implications of TUBB8 mutations
Previous analysis based on the known atomic structure of the α/β tubulin heterodimer (PDB: 3JAS)41 implied that the S176L mutation may result in the disruption of longitudinal interactions between heterodimers in the microtubule lattice and thereby affect microtubule stability.40 The p.T143Dfs*12 mutation will result in a truncated protein (154 N-terminal amino acids rather than the full-length 444 amino acid polypeptide) that cannot be incorporated into heterodimers. We mapped the remaining affected TUBB8 residues onto the same tubulin atomic structure. As shown in online supplementary figure S2, T285 is located within the M-loop, which is essential for lateral interactions within the microtubule. I210, N348, V255 and T238 are buried within the β-tubulin structure. Their mutation could affect folding and stability. Residues 27–33 may also contribute to β-tubulin stability. Amino acid R262 may form a salt bridge with D417, which is involved in kinesin binding;42 the R262W mutation may therefore weaken this interaction. In summary, the newly discovered TUBB8 mutations described here have the potential to interfere with microtubule function via tubulin deficit, to cause defects in microtubule behaviour by affecting β-tubulin folding, heterodimer assembly, or tubulin dynamics (via interfering with lateral contacts within the microtubule lattice), or to affect interaction with kinesin and perhaps other microtubule-associated proteins.
α/β heterodimer assembly in vitro and effect of mutant TUBB8 expression on microtubule architecture in cultured cells
To investigate whether the newly discovered mutations in TUBB8 indeed cause defects in β-tubulin folding and α/β heterodimer formation, we followed coupled transcription/translation reactions kinetically. This allowed us to monitor the transit of newly synthesised polypeptides through the various β-tubulin/chaperone complexes that contribute to the chaperone-dependent assembly pathway.43 None of the mutations had a substantial influence on translational efficiency (figure 4A); the minor band migrating at about 50 kDa (just below the major full-length product at 55 kDa and present in all the TUBB8 sequences analysed with the exception of the T143D*fs mutation, which produces a prematurely terminated translation product migrating at about 16 kDa) is probably a product of internal translational initiation. On the other hand, most of the TUBB8 missense mutants displayed a spectrum of differences compared with the wild-type control, including in many cases a reduction in the yield of de novo assembled heterodimers that could result in tubulin haploinsufficiency. This is particularly evident in the case of I120V, T238M and R262W (figure 4B). For the p.T143Dfs*12 and p.(E27_A33del) mutations, there is either a complete absence of folding intermediates (in the case of p.T143Dfs*12), or the generation of only a binary complex with the cytoplasmic chaperonin, CCT32 (in the case of p.(E27_A33del)). As expected, in neither case are de novo synthesised polypeptides containing either of these mutations capable of assembly into native α/β heterodimers. We conclude that both these mutations can be considered as functionally null.
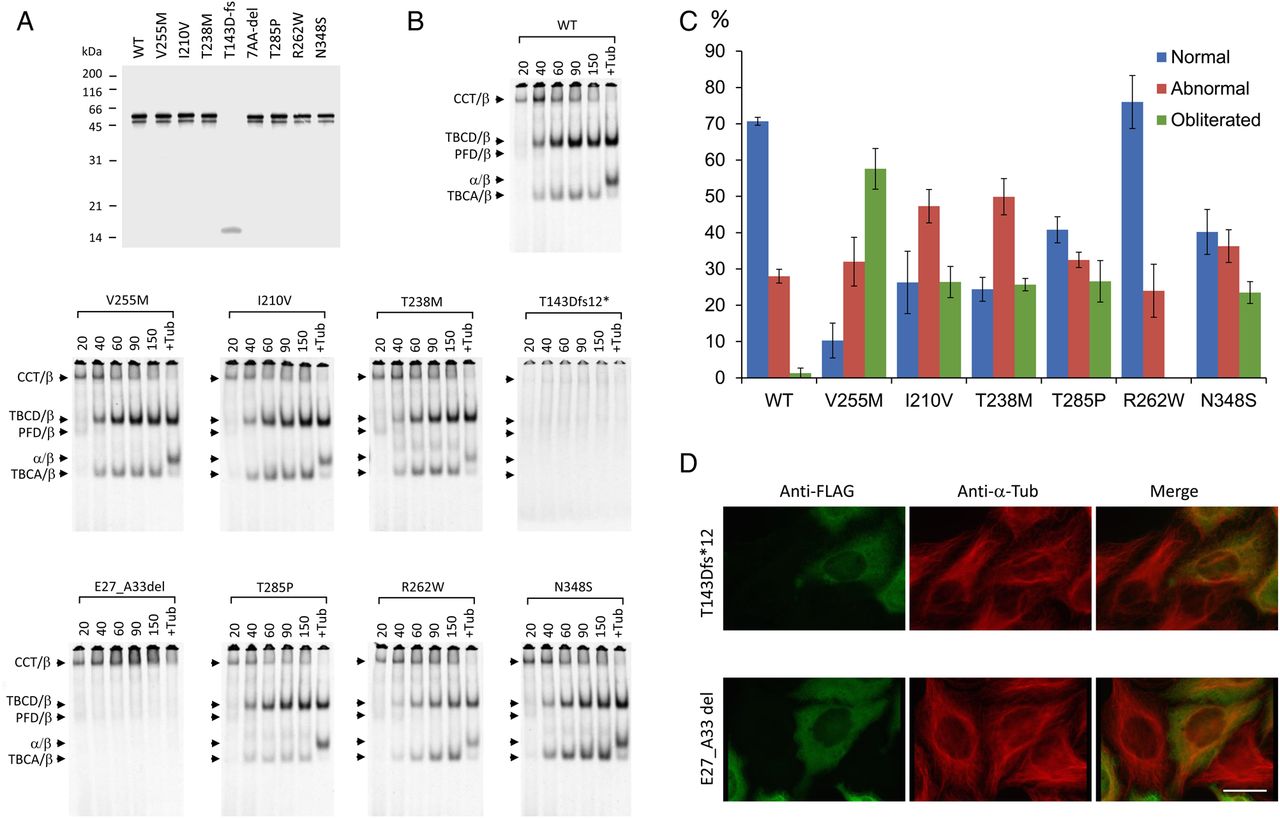
Kinetics of heterodimer assembly in vitro and microtubule phenotypes resulting from expression of wild-type and mutant forms of TUBB8 in cultured cells. (A) Analysis by 12% SDS-PAGE of the products of 35S-methionine-labelled transcription-translation reactions driven by plasmids encoding wild type and mutant forms of TUBB8. (B) Analysis on 4.5% native polyacrylamide gels of the products of transcription-translation reactions done in an identical manner to those shown in (A) for the times shown (in minutes) above each panel. Arrows (top to bottom) show the migration positions of the binary complex formed between newly translated polypeptides and the cytosolic chaperonin, CCT (CCT/β), the TBCD/β-tubulin complex, the Prefoldin/β-tubulin complex, the native α/β tubulin heterodimer and the TBCA/β-tubulin complex, respectively.32 (C) Quantitative analysis of microtubule phenotypes in HeLa cells expressing high levels of FLAG-tagged constructs encoding the mutations shown. The phenotypes (characterised as normal, abnormal or obliterated: examples shown in online supplementary figure S3) were analysed and scored as described previously.40 About 200 transfected cells were assessed for each category in each of three independent experiments. The figure shows mean percentages (±SD) of cells expressing wild-type or mutant TUBB8 and assigned to each category. (D) Expression of the T143Dfs*12 and E27_A33del TUBB8 mutations. HeLa cells transfected with constructs engineered for the expression of the truncation and seven amino acid internally deleted mutant forms of TUBB8, each bearing an in-frame C-terminal FLAG tag, were examined by immunofluorescence microscopy using an anti-FLAG antibody (to detect the transgene, shown in green) and an anti-α-tubulin antibody (to detect the endogenous microtubule network, shown in red). Note the appearance of the transgene as a diffuse mottled stain throughout the cytoplasm in each case, with no evidence of co-assembly into microtubules. Bar=10 µm.
To investigate the effect of the various TUBB8 mutants on microtubule behaviour, we transfected FLAG-tagged constructs into cultured HeLa cells. Consistent with our previously described analysis of missense mutations in TUBB8,40 most of the newly discovered missense mutations had a deleterious effect on the organisation of cytoplasmic microtubules (see online supplementary figure S3). A quantitative analysis of the microtubule phenotypes observed upon expression of wild-type and missense TUBB8 mutations showed that, with the exception of R262W, all had a greater propensity than the wild-type protein to cause microtubule obliteration (figure 4C). In contrast, cells transfected with either of the two functionally null mutations expressed the transgene in a diffuse mottled pattern distributed throughout the cytoplasm, with little or no effect on the organisation of the endogenous microtubule network (figure 4D). Given that the deletions in question each result in the excision of a critical portion of the β-tubulin polypeptide, we conclude that expression of these mutant proteins results in their deposition as insoluble cytoplasmic aggregates. The R262Q mutation was previously reported to cause a substantially diminished yield of properly assembled α/β-tubulin heterodimers in vitro, but had a relatively modest effect on the microtubule network upon expression in cultured cells.40 Notably, a different mutation at the same locus, R262W, also resulted in a reduced yield of assembled heterodimers (figure 4B). However, similar to cells transfected with wild-type, cells transfected with R262W had microtubules with a generally normal appearance (figure 4C). Thus, as would be expected, different amino acid substitutions at the same locus can cause different effects and result in variable phenotypes.
TUBB8 mutations and spindle assembly
To establish a causal relationship between mutations and phenotypes, we microinjected wild-type and mutant TUBB8-encoding RNAs into mouse oocytes Although mice (in common with all other non-primate species) do not harbour a gene encoding TUBB8, our previous work showed that expression of wild-type TUBB8 induced in mouse oocytes as a result of microinjection of recombinant RNA does result in co-assembly of TUBB8-containing heterodimers into meiotic spindle microtubules, thus setting a precedent for the usefulness of this approach.40 Compared with the appearance of uninjected control oocytes, oocytes microinjected with wild-type TUBB8 RNA had a spindle with a less dense appearance (figure 5), reinforcing our inference that β-tubulin isotype composition itself influences spindle morphology. In addition, expression of microinjected RNA encoding I210V, T238M, V255M, T285P and N348S TUBB8 in mouse oocytes resulted in completely or severely impaired spindle assembly (figure 5A,B), precisely recapitulating the spindle status in the corresponding patient oocytes. The absence of an anti-FLAG signal in the spindles of oocytes microinjected with RNA encoding p.(E27_A33del) confirmed that the effect of this mutation is equivalent to a null. Microinjection of R262W RNA resulted in a normal spindle (figure 5), consistent with the observation that expression of this mutant TUBB8 in HeLa cells does not disrupt microtubule architecture. In this respect, R262W is unique among known TUBB8 missense mutations.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
TUBB8 RNA microinjection, spindle morphology and assembly. (A and B) Immunostaining of mouse oocytes 12 hours after germinal vesicle breakdown (GVBD). GV oocytes were injected with a high (1000 ng/μL) concentration of either wild type (n=143) or mutant (I210V (n=67), T238M (n=53), R262W (n=66) and E27_A33del (n=81), (panel A) V255M (n=58), T285P (n=50) or N348S (n=57), (panel B) RNA encoding C-terminally FLAG-tagged TUBB8. Normal mouse GV oocytes (n=131) without injection were cultivated in vitro for maturation. In each case, the oocytes were stained to visualise chromosomes (Hoechst 33342; blue), spindles (ß-tubulin; green) and TUBB8 (FLAG; red) and examined by confocal microscopy.
Discussion
In this study, we identified eight novel heterozygous/homozygous mutations and one recurrent de novo mutation in TUBB8 from nine independent families with infertile females (figure 1 and online supplementary figure S1). Seven are heterozgyous missense, while two are homozygous mutations resulting in the expression of either a polypeptide with an internal deletion of seven amino acids or a truncated protein caused by a nucleotide insertion mutation and containing only 154 β-tubulin N-terminally derived amino acids. Taken together with our previous analysis,40 we have now identified 16 patients carrying a mutation in TUBB8 out of a total of 43 infertile women (ie, 36.3%) with MI oocyte arrest. It follows that other as yet unknown genetic defects can also contribute to infertility caused by oocyte maturation arrest. The discovery of the patients described here extends the spectrum of phenotypes in oocytes with meiotic arrest as a result of mutation in TUBB8 in two important respects. First, some morphologically MII oocytes can be fertilised, but the ensuing embryos become arrested at an early developmental stage, and second, some MI oocytes have a visible spindle (figures 2 and 3). Among missense mutations, most are predicted to affect microtubule behaviour by impacting either β-tubulin stability or lateral contacts between protofilaments (see online supplementary figure S2). Indeed, many of these were found to affect α/β heterodimer assembly in vitro, particularly with respect to the yield of material migrating as native heterodimer (figure 4B). Moreover, in several cases they disrupted microtubule organisation upon expression in cultured cells (figure 4D) and interfered with proper spindle assembly upon expression in mouse oocytes (figure 5).
We highlighted the location of all mutations identified in this and our previous study40 in the sequence of TUBB8 (see online supplementary figure S1A). Most are located in exon 4. Our previous analyses showed that various TUBB8 missense mutations resulted in impaired microtubule behaviour—including proper spindle assembly—by dominant negative effects.40 This mechanism also applies to some of the newly discovered heterozygous missense mutations reported here. However, in at least one case (R262W), the mutation had no discernable effect on microtubule architecture in cultured cells or on spindle assembly upon expression in mouse oocytes. The R262W mutation therefore seems more likely to exert its effect either via haploinsufficiency (reflected in the reduced yield of native tubulin heterodimers generated in in vitro folding assays, figure 4B) or as a result of one or more predicted alterations in the binding of kinesin (see online supplementary figure S2) rather than a dominant negative effect on microtubule stability or dynamics (figure 5B). On the other hand, oocyte maturation arrest can also be caused by homozygous mutations in TUBB8 (T143Dfs*12 and E27_A33del), each with a recessive inheritance pattern. We were surprised to find spindle structures in oocytes carrying these mutations since they both render the expressed protein functionally null (see online supplementary figure S4A–C). At least in the latter case, a defective spindle was observed (figure 2I). It therefore seems probable that spindle microtubules in these homozygous mutant-bearing oocytes are polymerised from pre-existing non-TUBB8-containing tubulin heterodimers expressed at an earlier developmental stage, without compensatory upregulated expression of other β-tubulin isotypes. Because patient oocytes were not readily obtainable, the latter conclusion was based on mRNA-based analysis from a single oocyte. Nonetheless, we interpret the spindle defects resulting from this mutation as reflecting an essential contribution of heterodimers containing TUBB8 to proper spindle structure in primates. Consistent with this inference, the spindles of mouse oocytes microinjected with wild-type TUBB8 RNA were morphologically distinct from uninjected controls (figure 5). We conclude that TUBB8 has a key role in determining the specific morphology and normal function of the primate meiotic spindle, and that the class of genetically determined diseases known as the tubulinopathies37 ,38 must now be extended to include female infertility caused by mutations in TUBB8.
Because of the scarcity of human oocytes available for experimental purposes, we were constrained in the number of these cells we could use for analysis by immunostaining. Nonetheless, our data are sufficiently compelling to justify the phenotypic classification of oocytes harbouring the TUBB8 mutations we describe into three distinct classes: MI oocytes with an impaired spindle (heterozygous mutations: S176L, T238M, V255M, T285P, N348S); MI oocytes that are dysfunctional but that have a visible but morphologically defective spindle (homozygous mutations p.T143Dfs*12 and p.(E27_A33del)) and morphological MII oocytes that either lack an identifiable spindle or contain an impaired spindle (I210V, T238M and N348S). Some of the oocytes morphologically identifiable as MII but with an impaired spindle can be fertilised and cleave, but these embryos later become developmentally arrested. Typically, in the context of clinical IVF/ICSI, only morphological MII oocytes identified by light microscopy are selected for fertilisation. In view of our discovery that oocytes morphologically recognisable as MII can be defective, determination of genetic variations in TUBB8 is likely to prove useful as a future additional criterion for evaluating the quality of patients' MII oocytes. We note that Dokshin et al44 demonstrated that stra8-deficient mouse oocytes can produce a polar body without premeitoic chromosomal replication or recombination, suggesting that oocyte growth and differentiation are genetically dissociable from meiosis in mice. Our observations provide direct evidence that as in mouse oocytes, meiosis and oocyte differentiation make independent contributions to the generation of a functional human egg.
Acknowledgments
The authors thank Dr Eva Nogales and Dr Rui Zhang for great help on structural analysis of TUBB8 mutations. The authors also thank Ke Qiao from Key Laboratory of Medical Molecular Virology, Ministry of Education and Public Health, School of Basic Medical Sciences, Fudan University for help with technical expertise in confocal microscopy.
References
Footnotes
NJC and YK are senior contributing authors.
Contributors RF, ZY, BL, MY, QS and GT contributed equally NJC, GT and LW conceived and designed the experiments. MY, ZY, XS, YK and BL collected the samples. RF, QS, GT, YX, BC, RQ performed the experiments and analysed the data. ZS, LJ and LH analysed the data. NJC and LW wrote the paper. All authors reviewed the manuscript.
Funding This work was supported by the National Basic Research Program of China (2015CB943300), the National Natural Science Foundation of China (81270747, 81270749, 81571397, 81571501, 81300485, 81300550), the 111 Project (B13016) and by a National Institutes of Health grant (5R01GM097376 to Dr Nicholas Cowan).
Competing interests None declared.
Patient consent Obtained.
Ethics approval Fudan University Institutional Medical Review Board, Shanghai, China. Reproductive Study Ethics Committee, the Ninth Hospital affiliated with Shanghai Jiao Tong University, Shanghai, China.
Provenance and peer review Not commissioned; externally peer reviewed.