Article Text

Abstract
Background and methods: Ring chromosomes are often associated with abnormal phenotypes because of loss of genomic material at one or both ends. In some cases no deletion has been detected and the abnormal phenotype has been attributed to mitotic ring instability. We investigated 33 different ring chromosomes in patients with phenotypic abnormalities by array based comparative genomic hybridisation (CGH) and fluorescence in situ hybridisation (FISH).
Results: In seven cases we found not only the expected terminal deletion but also a contiguous duplication. FISH analysis in some of these cases demonstrated that the duplication was inverted. Thus these ring chromosomes derived through a classical inv dup del rearrangement consisting of a deletion and an inverted duplication.
Discussion: Inv dup del rearrangements have been reported for several chromosomes, but hardly ever in ring chromosomes. Our findings highlight a new mechanism for the formation of some ring chromosomes and show that inv dup del rearrangements may be stabilised not only through telomere healing and telomere capture but also through circularisation. This type of mechanism must be kept in mind when evaluating possible genotype–phenotype correlations in ring chromosomes since in these cases: (1) the deletion may be larger or smaller than first estimated based on the size of the ring, with a different impact on the phenotype; and (2) the associated duplication will in general cause further phenotypic anomalies and might confuse the genotype–phenotype correlation. Moreover, these findings explain some phenotypic peculiarities which previously were attributed to a wide phenotypic variation or hidden mosaicism related to the instability of the ring.
Statistics from Altmetric.com
Ring chromosomes are usually associated with abnormal phenotypes due to the loss of material at both or at least one chromosome end. Thus, in principle, the abnormal phenotypes are essentially due to haploinsufficiency of those dosage sensitive genes contained in the deleted segment(s). The finding of ring chromosomes without apparent loss of genetic material in subjects with abnormal phenotypes led investigators to hypothesise that the ring formation with the related difficulties in the sister chromatid separation at cell division induced the generation of secondary aneuploid cells. Some aneuploidies, being lethal at the cellular level, would in turn give rise to increased cell death rate. Altogether this situation should lead to the “ring syndrome”1 that in cases with intact ring chromosomes is characterised, independently of the chromosome involved, by severe growth failure, minor dysmorphic features, and mild to moderate mental retardation, without major malformations. In a review of 207 cases, Kosztolányi2 estimated that one fifth of subjects with autosomal rings are affected by the “ring syndrome” phenotype. Indeed, more recent papers have demonstrated that intact ring chromosomes may cause areas of hypopigmentation along the lines of Blaschko as the only sign of ring-induced mosaicism,3 or specific features such as a characteristic type of epilepsy and electroencephalographic pattern as reported for several ring (20) chromosomes,4 thus weakening the hypothesis of the “ring syndrome”. Moreover, fluorescent in situ hybridisation (FISH) analysis at first, and more recently whole genome array screenings, have demonstrated that in most of the cases a cryptic deletion is at the basis of the phenotypic abnormalities in apparently intact rings.5–7 Recently the case of an r(14) has been reported with the combination of an inverted duplication with a terminal deletion characterised using high resolution molecular karyotyping and FISH.8 The patient presented overlapping clinical features described in terminal deletion, duplication and ring chromosome 14 cases. By examining 33 probands with ring chromosomes through array based comparative genomic hybridisation (CGH) we detected the same situation in seven of them, suggesting both a new mechanism of ring formation and a warning for clinical geneticists to consider this possibility while performing genotype–phenotype correlations.
MATERIALS AND METHODS
Patients
The initial chromosome examination which detected a ring chromosome was performed in 17 cases in different cytogenetic laboratories in Italy and in 16 cases in Zurich, Switzerland. In one case (case 33, from Zurich), the ring chromosome was transmitted from a mildly affected mother with low mosaicism to a non-mosaic daughter, while in all the other cases the ring formation had occurred de novo. Three cases, 8 (case 14 in Ballarati et al9), 9 (case 10 in Ballarati et al9 and Guala et al10) and 25 (Baumer et al11) had already been published.
Array CGH
In 28 cases array based CGH was performed using the Agilent Human Genome CGH Microarray Kit 44B (Agilent Technologies, Santa Clara, California, USA), and in six cases the 244A Kit. In one case (case 21) we used both platforms. These platforms are high resolution oligonucleotide based microarrays that allow genome-wide survey and molecular profiling of genomic aberrations with a resolution of about 100 kb and 20 kb, respectively. Labelling and hybridisation were performed following the protocols provided by Agilent. Briefly, 500 ng of purified DNA of a patient and of a control (Promega Corporation, Madison, Wisconsin, USA) were double-digested with RsaI and AluI for 2 h at 37°C. After 20 min at 65°C, DNA of each digested sample was labelled, by the Agilent random primers labelling kit, for 2 h using Cy5-dUTP for the patient DNA and Cy3-dUTP for the control DNA. Labelled products were column purified and prepared according to the Agilent protocol. After probe denaturation and pre-annealing with 50 μg of Cot-1 DNA (Invitrogen, Carlsbad, California, USA), hybridisation was performed at 65°C with rotation for 40 h. After two washing steps the arrays were analysed with the Agilent scanner and the Feature Extraction software (v8.0; v9.1.3). Graphical overview was obtained using the CGH analytics software (v3.1; v3.4). All the breakpoint sequences were analysed with self chain and segmental duplication repeats tools from UCSC (http://genome.ucsc.edu/).
FISH
To confirm array CGH results and demonstrate the inversion of the duplication in patients 7 and 13, FISH experiments were performed. All bacterial artificial chromosome (BAC) probes used were selected according to the UCSC Human Genome Browser and were obtained from the human library RPCI-11. DNA extraction and FISH experiments were carried out as previously described.12 The inversion of the duplication was determined in two cases by dual colour FISH experiments according to standard protocols.
Genotyping
Genotyping of polymorphic loci was performed by amplification with primers labelled with fluorescent probes (ABI 5-Fam and Hex) followed by analysis on an ABI 310 Genetic Analyzer (Applied Biosystems, Foster, California, USA). The UCSC Genome Browser maps and sequences were used as references. Amplification was performed with Taq Gold (Applied Biosystems) using standard protocols. In case 13, primers were designed in the duplicated region. The sequences of the informative markers are the following:
15DUP3 left: CTCAGCGAGAGACCATCCTC, right: AAGAGAGCAGGAGGGAAAGC;
15DUP4 left: CCAAAAAGTGCATGGTGTTG, right: AGGGAATGTGACCTGTGGAG;
15DUP11 left: CAGAATTCGGGACAGGAAAA, right: CCAATCGCCTGCTCTCTTAG;
15DUP13 left: TGGAGAACACCTGGAAAAGG, right: CGGCCGTTCTCATGTATCTT;
15DUP15 left: TAAGCAAGCACCAAGCACAC, right: TTGTTGCTTCTGTGGGAAAA
RESULTS
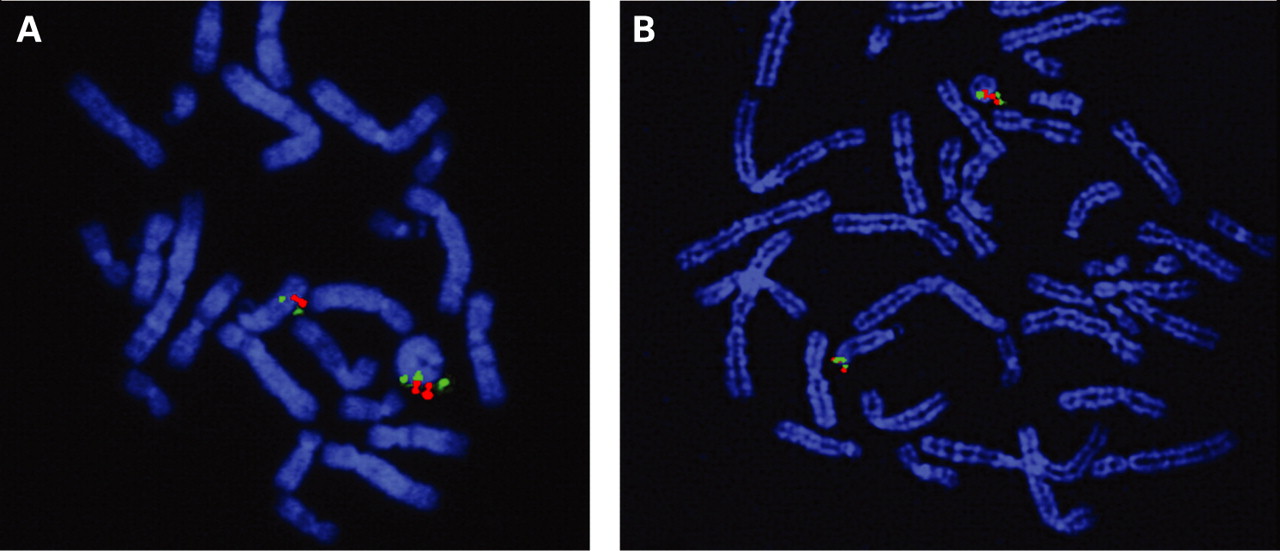
We identified seven dup del or inv dup del rearrangements among 33 different ring chromosomes (fig 1). The denomination of “inv dup del” was reserved to those two cases (cases 7 and 13) in which, thanks to the availability of lymphoblastoid cell lines, we demonstrated by dual colour FISH that the duplication was inverted. In case 7 the BACs used were RP11-164b1 (biotin labelled) and RP11-122a8 (digoxigenin labelled) (fig 2A); in case 13 the inversion was identified by BAC RP11- 1072p10 (biotin labelled) and RP11-522b15 (digoxigenin labelled) (fig 2B). The orientation of the duplication could not be investigated in the five cases with a “dup del” rearrangement (cases 5, 10, 21, 26, 33). The chromosomes involved were: chromosome 13 (cases 5, 7 and 10), chromosome 15 (case 13), chromosome 18 (cases 21 and 26) and chromosome 22 (case 33).


Data on array-CGH results and parental origin of the 33 cases are given in table 1.
The clinical features of the seven patients with inv dup del or dup del ring chromosomes and the array CGH findings with the resulting genomic imbalance are listed in table 2.
In the two inv dup del cases (cases 7 and 13) microsatellite analysis in patients and parents with markers mapping to the deleted regions (D13S173, D13S1315: case 7; D15S642, D15S107: case 13) indicated a paternal origin of the rearrangement in both cases (fig 3). Moreover, microsatellite markers of the duplicated region showed that the rearrangement was intrachromosomal since only two alleles, the paternal one showing a double peak area, had been detected for all the informative markers (D13S1283, D13S790, D13S1323; 15DUP3, 15DUP4, 15DUP11, 15DUP13, 15DUP15) (fig 3A,B).

{kind=link}
{kind=link}
{kind=link}
In case 33, with a dup del(22q), the rearrangement was probably maternal in origin. We did not perform microsatellite analysis since parental DNA was not available. However, the mother who had some phenotypic abnormalities showed a low mosaicism (1 out of 50) for the same ring in lymphocytes.
In case 21 with an inv dup del(18p), the array CGH results with the Agilent Kit 44B (100 kb resolution) identified in 18p a single deleted oligonucleotide (170,229–170,285 kb). A second analysis with the Agilent Kit 244A, with a resolution of about 20 kb, confirmed a real terminal deletion of 207.5 kb (fig 1F). In this case, as in case 26, a second deletion was detected at 18q.
DISCUSSION
Ring formation in the seven dup del/inv dup del rearrangements
The classic mode of formation of ring chromosomes is breakage in both arms of a chromosome, loss of distal fragments followed by fusion of the proximal broken ends. In this case the patient has a partial monosomy for the distal short arm and distal long arm. In other cases, especially patients with no or very mild abnormal phenotype, loss of the telomeres with retention of subtelomeric sequences has been reported. It has been assumed that even this small genetic loss may contribute to an abnormal phenotype (patient 2 in Sigurdardottir et al13). The other mode of formation is telomere-to-telomere fusion, with telomeric and subtelomeric sequences being retained, and in this case no genetic material would be lost. It has been hypothesised that the phenotypic abnormalities in this type of ring chromosome are due essentially to what is called “dynamic mosaicism” with pre- and postnatal growth retardation and microcephaly as consistent features.2 14 Several examples show the presence of specific phenotypes otherwise typically associated with dominant gene mutations in patients carrying ring chromosomes not deleted for the responsible genes.15 16 Thus, hemizygosity of the responsible gene(s) in a significant proportion of somatic cells due to the ring instability may explain the phenotype.
Obviously, cryptic deletions of critical sequences distal to the commercial subtelomeric clone in one of the two arms had also been suspected to be responsible for some phenotypic abnormalities.17
Our study demonstrates that some ring chromosomes have been formed through a more complex rearrangement leading to concurrent deletion and contiguous duplication. Although the inversion of the duplicated region has been demonstrated only in two cases (cases 7 and 13) in whom lymphoblastoid cell lines were available, we assume that the duplication was inverted also in all the other ring cases with a duplication contiguous to a terminal deletion. Thus, the rings sharing the same type of rearrangement were 7 out of 33. This suggests a different mechanism at the basis of such ring chromosomes with the formation of an intermediate inv dup del chromosome that circularises to stabilise itself. Obviously this mechanism has important implications for the phenotype which is not only due to the simple deletion as generally assumed for ring chromosomes but also to the associated duplication.
A single case of a ring chromosome 14 containing a terminal deletion and an additional inverted duplication proximal to the deletion has been recently reported.8 There are a few reports in the literature, suggesting similar rearrangements. One case is a ring chromosome 9 with an inverted duplication of the distal segment of 9p.18 However, in this case the inverted duplication was only suggested by high resolution banding, and there was no apparent concomitant distal 9p deletion. Another case is an r(21) with partial trisomy for much of 21q and partial monosomy for distal 21q (patient 10 in McGinniss et al19) found in a patient with mild Down stigmata. The molecular and cytogenetic data of this patient were consistent with the model of r(21) formation in which asymmetric breakage and reunion of the long arms of an intermediate isochromosome or robertsonian translocation chromosome generated a large r(21). An unusual r(21) in mosaic state, in a child who had some manifestations of Down syndrome, has been reported by Palmer.20 Molecular studies demonstrated that the ring carried a duplicated region and a concomitant distal deletion and suggested the following mechanism: an initial robertsonian translocation occurred between one paternal and one maternal chromosome 21 in a trisomic cell, followed by distal breakage and deletion in both long arms of the translocation, leading to the formation of a ring chromosome. Three older reports described four mosaic cases each involving two cell lines, one with an isochromosome or robertsonian translocation chromosome and the other with a ring chromosome.21–23 In these cases the authors suggested a post-zygotic secondary event in the translocation chromosome to explain the ring formation.
Our findings indicate a different mechanism of formation of the deleted and duplicated ring chromosome. In fact, the duplicated region never involved the proximal portion of 13q, 15q, 18p, 22q as expected in an isochromosome or a robertsonian chromosome but rather involved more distal regions contiguous to the deletion region. The most well known mechanism for formation of inv dup del type rearrangements is non-allelic homologous recombination between homologous segmental duplications located at the rearrangement’s breakpoints,24–26 leading to an intermediate dicentric chromosome. In our cases, the cytogenetic breakpoints do not fall in regions known to contain segmental duplications; therefore, the formation of the dicentric chromosomes cannot be explained by non-allelic homologous recombination (NAHR). The resolution of the array platforms used is too low to reveal if specific DNA motifs leading to instability such as palindromic AT-rich repeats27 or other non-B DNA structures28 are present at the rearrangements’ breakpoints. Moreover, results from microsatellite marker examinations of the duplicated region in one r(13) (case 7) and in one r(15) (case 13) suggest that the rearrangement is intrachromosomic, indicating that at least in these cases non-homologous end joining can be responsible for the formation of the dicentric, as demonstrated initially in maize29 30 and more recently in humans—for example, in some cases of inv dup del(1p).31
An asymmetric breakage of the dicentric would lead to two different abnormal chromosomes, one inv dup del chromosome and a simply deleted one, that could be stabilised by different mechanisms. Telomere healing and telomere capture are the most well known mechanisms to stabilise broken chromosomes and there are many descriptions in the literature of how these work.32–35 In our cases the stabilisation of the inv dup del chromosome is achieved through the formation of the ring, representing a new mechanism through which such inv dup del chromosome can stabilise itself. According to our hypothesis the mechanism leading to dup del rings is a multi-step process: a prezygotic event leading to the formation of the dicentric, a postzygotic breakage leading to the inv dup del chromosome, and the circularisation of the inv dup del in some cells. The finding that these rings are present in most of the cells and no cell lines showing either the inv dup del or the simply deleted chromosome have been found, can be explained assuming an early selection versus the most viable cell line(s). In this case an important selection factor may be the restriction of telomerase expression to the first embryonic stages36 impairing the broken chromosomes to be stabilised by telomere healing. An example showing that post-zygotic events may produce a mosaicism present in the fetus but not in the adult is provided by the classical inv dup del(8p). We37 and others38 demonstrated that it can be found in mosaic with a del(8p) during the fetal life, although in the great majority of postnatal cases the inv dup del(8p) is present in all cells.
Ring formation in the other cases
In the other 26 cases, array CGH identified the following rearrangements.
In 18 cases a distal deletion in one arm was present. For the 16 cases involving acrocentric chromosomes (chromosomes 13, 15 and 22) a second deletion at the short arm is likely and cannot be excluded since these regions are not represented in the array platforms. No recurrent breakpoints had been found in rings involving the same chromosome and none of the breakpoints was characterised by the presence of segmental duplications. In cases 1 and 2, concerning an r(4) and an r(5), a single deletion at the level of the short and long arm respectively had been detected. Although a second very small deletion at the opposite arm cannot be excluded while using higher resolution array platforms, it is possible that ring chromosome formation in these cases acted as an alternative way to stabilise a broken chromosome next to telomere healing and capture. In three cases of r(18) (cases 22, 24 and 25) array CGH identified two distal deletions in both arms. A similar situation in two ring chromosomes 18 characterised by FISH and microsatellite analysis has been described (cases 4 and 5 in Stankiewicz et al5). In these cases the ring chromosome could have been formed through a classical mechanism involving two breakages in both arms, loss of the distal fragments followed by fusion of the proximal broken ends. In case 23, with an r(18), the whole 18p arm was duplicated and a small distal deletion was present at the opposite end. The phenotype was mild, according to duplication of the whole 18p. Concerning the rearrangement, we cannot exclude a small distal deletion in 18p with a similar mechanism as demonstrated in other inv dup del rearrangements; alternatively, a pericentric inversion of chromosome 18 preceding the ring formation or an 18p direct/inverted duplication could be possible. A further blood sample from the patient and further analysis are necessary to delineate the real situation.
In three patients with epilepsy and a ring chromosome 20 in mosaic state (30–40%) (cases 27–29), we could not identify any genomic loss even using the platform at about 20 kb resolution. Many other similar cases have been reported,4 14 39 and the ring in these cases might be derived from telomere–telomere fusion, as already demonstrated by quantitative FISH analysis in one mosaic r(20) patient.4
In case 30, with a ring chromosome 21 in a patient with Down syndrome, array CGH identified an interstitial duplication followed by a distal triplication; a distal 21q deletion could not be shown.
Genotype–phenotype correlations
The findings that 21% of the ring chromosomes we studied show not only the expected deletion(s) but also a contiguous duplication have important genotype–phenotype implications. It is essential to emphasise that, at the beginning, the identification of two out of seven inv dup del rearrangements in ring chromosomes was absolutely fortuitous. Cases 7 and 13 belong to two series of patients, with deletions of chromosome 13 and chromosome 15, respectively, that were analysed by array CGH and FISH for genotype–phenotype correlation studies. These unexpected findings prompted the investigations of further cases.
It is obvious that in a ring chromosome, phenotypic correlations cannot be done any more assuming a simple deletion before having excluded additional duplicated segments. So far, it had been assumed that a ring of the length of the normal homologue has lost little material, and thus the adverse consequences for the phenotype are minor, and vice versa. This would obviously not be true if a relatively large ring has a deletion plus a duplication which compensate each other with respect to size alteration. We cannot extrapolate from our dup del ring cases which specific traits were due to the deletion and which ones to the duplication: some of the patients were very young at clinical examination and several of them have been examined by different clinicians. However, at least in case 5, with the r(13) having a deletion and a duplication of approximately 6 Mb each, oligohydramnios and cystic kidney may be attributed to trisomy 1340 41 and not to monosomy for the distal 13q33.3-qter region.9 In case 13 with the r(15) having a deletion of 5 Mb and a duplication of 2 Mb, abnormal hand ossification with an additional phalanx on the third finger was observed. Similar findings have been reported in an old case with partial trisomy 15q42 but never in cases with a simple 15q deletions.7 41 43 Thus, if up to now the explanation for the spectrum and severity of symptoms in patients with apparently identical ring chromosomes was attributed to mitotic instability of the ring, causing the formation, in a mosaic state, of secondary rings with either larger deletion or additional duplication or both, from now on the possibility that the ring is not only deleted but also duplicated should also be taken into consideration.
In the other ring cases where the expected deletion had been confirmed by array CGH the phenotype was more or less severe according to the extension of the deleted region. For example, case 9 with a 13q deletion of 33.7 Mb died at the age of 3 years because of severe congenital anomalies,10 whereas case 8 with a 13q deletion of 4.5 Mb has moderate mental retardation but no malformations (cases 10 and 14 in Ballarati et al9).
In case 30 segmental triplication of chromosome 21 could be demonstrated. Previous assumptions of mosaic tetrasomy of 21 turned out to be erroneous: these cases with mosaicism for a metacentric extra chromosome turned out to be examples of mosaic tetrasomy 12p—the Pallister-Killian syndrome.41 The 11.2 Mb duplication in our patient covers the 21q21.3-q22.2 region considered to be most important for Down syndrome phenotype, hence explaining her Down syndrome phenotype. We have recently observed another case with the phenotype of mild and atypical Down syndrome in whom array CGH results demonstrated a complex rearrangement of chromosome 21 with deleted and duplicated and also one triplicated segment (AS, unpublished observation). The reason why such cases were not detected before is probably that, unless there is an extra chromosome, these cases would not be investigated for potential segmental triplication with molecular methods; array-CGH, however, shows the triplication without special focused examinations.
Conclusions
We consider that re-examination using array CGH will solve any discrepancy in patients with a clinical phenotype not congruent with the size, the banding pattern of the ring and the FISH results. Array CGH can show additional aneuploid segments, either deletions or duplications or both. Therefore we suggest that all ring patients are (re-)examined using array CGH.
Acknowledgments
The authors are thankful to the patients and parents as well as the referring physicians for excellent cooperation.
REFERENCES
Footnotes
↵*The first two authors contributed equally to the work
Funding: This work was supported by PRIN 2005 and 2006 (to OZ and ER), Fondazione Mariani and Fondazione CARIPLO (all to OZ). MR and AS received a grant from the Swiss National Foundation (No.32-113635/1).
Competing interests: None declared.
Patient consent: Informed consent was obtained for the publications of the patients’ details in this article