Article Text

Abstract
Background Male infertility due to multiple morphological abnormalities of the sperm flagella (MMAF) is a genetically heterogeneous disorder. Previous studies revealed several MMAF-associated genes, which account for approximately 60% of human MMAF cases. The pathogenic mechanisms of MMAF remain to be illuminated.
Methods and results We conducted genetic analyses using whole-exome sequencing in 50 Han Chinese probands with MMAF. Two homozygous stop-gain variants (c.910C>T (p.Arg304*) and c.3400delA (p.Ile1134Serfs*13)) of the SPEF2 (sperm flagellar 2) gene were identified in two unrelated consanguineous families. Consistently, an Iranian subject from another cohort also carried a homozygous SPEF2 stop-gain variant (c.3240delT (p.Phe1080Leufs*2)). All these variants affected the long SPEF2 transcripts that are expressed in the testis and encode the IFT20 (intraflagellar transport 20) binding domain, important for sperm tail development. Notably, previous animal studies reported spontaneous mutations of SPEF2 causing sperm tail defects in bulls and pigs. Our further functional studies using immunofluorescence assays showed the absence or a remarkably reduced staining of SPEF2 and of the MMAF-associated CFAP69 protein in the spermatozoa from SPEF2-affected subjects.
Conclusions We identified SPEF2 as a novel gene for human MMAF across the populations. Functional analyses suggested that the deficiency of SPEF2 in the mutated subjects could alter the localisation of other axonemal proteins.
- spef2
- sperm
- infertility
- exome
- sequencing
Statistics from Altmetric.com
Introduction
Infertility has become one of the major issues of human health, affecting 10%–15% of the general population worldwide.1 Male factors are identified in approximately half of the infertile couple, and the diagnosis and treatment of male infertility therefore elicit an increasing interest. Notably, multiple morphological abnormalities of the sperm flagella (MMAF) have been known to affect sperm motility and cause male primary infertility in humans.2 Men with MMAF present sperm defects characterised by absent, short, bent, coiled and/or irregular flagella without any other symptoms of immotile-cilia syndrome.2
Autosomal recessive inheritance has been reported in human MMAF.2 Previous studies have shown that mutations in DNAH1 (MIM: 603332) are the most frequent genetic cause in human MMAF.2 Moreover, mutations in the genes encoding cilia and flagella-associated proteins, such as CFAP43 (MIM: 617558), CFAP44 (MIM: 617559), CFAP69 (MIM: 617949) and CFAP251 (MIM: 618146), can also cause male infertility with MMAF.3–7 Furthermore, several other genes are newly reported to be associated with MMAF, including AK7 (MIM: 615364), ARMC2, ARMC3 (MIM: 611226), CEP135 (MIM: 611423), FSIP2 (MIM: 615796) and QRICH2 (MIM: 618304).8–13 These findings demonstrated that MMAF is genetically heterogeneous. The pathogenic mechanisms in the remaining genetically unexplained MMAF cases are still unknown, and the potential involvement of new genetic factors remains to be illuminated. Here we conducted genetic analyses using whole-exome sequencing and identified three unrelated MMAF individuals (two Chinese and one Iranian) harbouring homozygous stop-gain mutations in SPEF2 ( sperm flagellar 2 ), indicating its important roles in sperm flagellar development and male fertility.
Materials and methods
Subjects and clinical investigation
In this study, 50 Han Chinese men with MMAF were recruited from The First Affiliated Hospital of Anhui Medical University (Hefei, China). Twenty-three of these subjects were enrolled from consanguineous families. A detailed history about the onset and progression of infertility was obtained from each participant. Peripheral whole blood samples were collected for genetic analyses. Moreover, the human exome data from a previously described cohort of 78 individuals with MMAF of North African, French and Iranian origins4 were reanalysed in this study. Clinical investigation revealed that all the probands in this study displayed a normal erection and ejaculation. These cases with primary infertility presented with MMAF phenotypes. Examinations revealed normal development of male external genitalia, normal bilateral testicular size, normal hormone levels and secondary sexual characteristics. The chromosomal karyotypes of all probands are normal (46; XY), and no microdeletion was found in the human Y chromosome. Moreover, primary ciliary dyskinesia (PCD)-associated symptoms (such as sinusitis, bronchitis, pneumonia and otitis media)14 were consulted very carefully and excluded from the MMAF cases by olfactory test with smelling bottles or chest X-ray scanning.
Whole-exome sequencing and bioinformatic analysis
Genomic DNAs were obtained from peripheral whole blood samples using the QIAamp Blood DNA Mini Kit (QIAGEN, Germany). The human exomes were enriched by the Agilent SureSelectXT Human All Exon Kit. Next-generation sequencing was conducted on the Illumina HiSeq X-TEN platform. The obtained raw reads were mapped to the human genome reference assembly (hg19) using the Burrows-Wheeler Aligner software.15 The Picard software was employed to remove PCR duplicates and evaluate the quality of variants. Then we investigated DNA sequence variants using the Genome Analysis Toolkit.16 The variants with read depths less than 4× were filtered out. Details of variant annotation can be found in the online supplementary methods. Details on the methods used for analysis of the 78 individuals with MMAF were described previously.4
Supplemental material
Semen characteristics and sperm morphological study
Fresh semen of human subjects was collected after 2–7 days of sexual abstinence and examined after liquefaction for 30 min at 37°C. Semen volume, sperm concentration and motility were evaluated in accordance with the WHO guidelines. Sperm morphology was analysed using H&E staining and differential interference contrast microscopy (DICM). For each subject, at least 200 spermatozoa were examined to evaluate the percentages of morphologically abnormal spermatozoa according to the WHO guidelines.
Transmission electron microscopy
For transmission electron microscopy (TEM), the prepared spermatozoa were immersed in 2.5% phosphate buffered glutaraldehyde, were washed with 0.1 mol/L phosphate buffer (PB, pH 7.2) three times, and were postfixed with 1% osmium tetroxide in 0.1 mol/L PB for 1–1.5 hours at 4°C. Dehydration was performed using 50%, 70%, 80%, 95% 100% ethanol and 100% acetone sequentially, followed by infiltration with 1:1 acetone and SPI-Chem resin (containing dodecenly succinic anhydride, N-methylacetamide, SPI-Pon 812 and DMP-30) overnight at 37°C. After infiltration and being embedded in Epon 812, ultrathin sections were stained with uranyl acetate and lead citrate, then were observed and photographed by TEM (TECNAI-10, Philips) with an accelerating voltage of 80 kV.
RT-PCR assays
The sperm samples were obtained from the SPEF2-mutated subject A028-II-1 and a healthy control man. The normal lung and brain samples were taken from the control subjects undergoing acquired lung bullae and brain trauma after obtaining informed consent. Total RNAs of different human tissues were extracted using the RNeasy Mini Kit (QIAGEN). Total RNAs of 1 µg were converted into cDNAs with SuperScript III Reverse Transcriptase (Invitrogen) and oligo (dT) primers (TaKaRa). The obtained cDNAs were used for subsequent PCR with transcript-specific primers (online supplementary table 1).
Immunofluorescence assays
Sperm cells were fixed in 4% paraformaldehyde and blocked in 20% donkey serum containing 0.4% (vol/vol) Triton X-100 before being incubated overnight at 4°C with the following primary antibodies: rabbit polyclonal anti-SPEF2 against peptides at 227-275aa (bs-11488R, Bioss, 1:100), anti-CFAP69 (bs-15278R, Bioss, 1:100) and monoclonal mouse anti-α-tubulin (T9026, Sigma, 1:500). Washes were performed using phosphate buffer saline, followed by 1-hour incubation at room temperature with highly cross-absorbed secondary antibodies (1:1000, dilutions: anti-rabbit-Alexa Fluor-488 for anti-CFAP69, anti-SPEF2; anti-mouse-Alexa Fluor-647 for anti-α-tubulin). Images were captured with a confocal microscope (Zeiss LSM 710).
Results
Among the cohort of 50 MMAF-affected Chinese men analysed by whole-exome sequencing in this study, two homozygous stop-gain variants in SPEF2 (MIM: 610172) were identified in unrelated probands from two consanguineous families (A028 and A042) (figure 1A). Both SPEF2 variants were confirmed by Sanger sequencing. The homozygous SPEF2 variant c.910C>T (p.Arg304*) in subject A028-II-1 was inherited from his heterozygous parental carriers (figure 1A). In the second family (A042) affected by SPEF2, the homozygous frameshift variant c.3400delA (p.Ile1134Serfs*13) was identified in subject A042-II-1 with MMAF (figure 1).
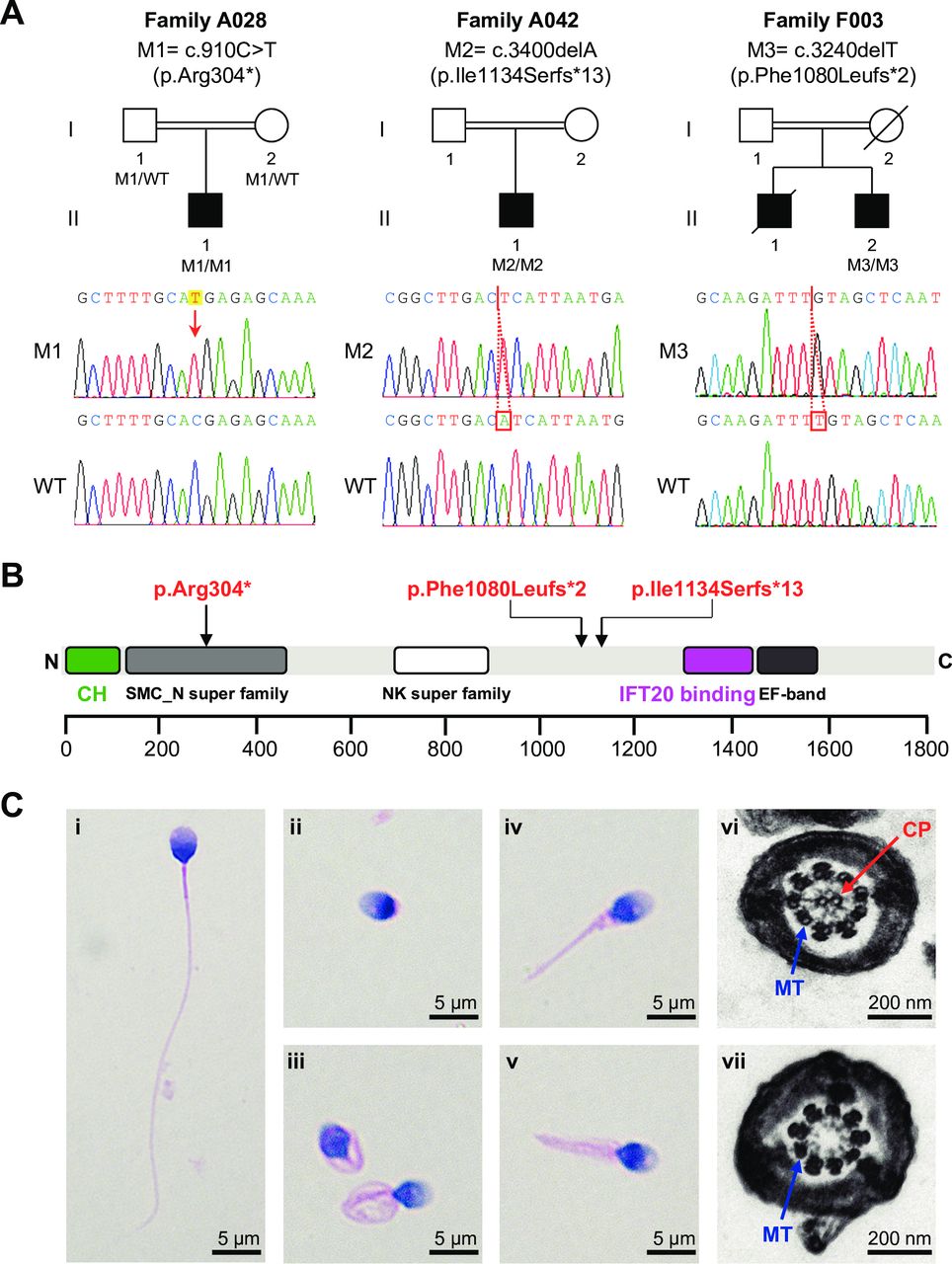
Homozygous stop-gain variants of SPEF2 and sperm morphological defects in the SPEF2-mutated probands with MMAF. (A) Pedigrees of the investigated families affected by homozygous SPEF2 variants. Sanger sequencing results are shown below the pedigrees. The variant positions are indicated by a red arrow or red boxes. (B) A schematic representation of SPEF2 protein and the variants identified in this study. The green box indicates the known calponin homology (CH) domain with a critical role in regulating ciliary function. The purple box indicates the IFT20 binding domain involved in sperm tail development.18 (C) Morphology and ultrastructure of spermatozoa from the SPEF2-mutated probands with MMAF. A normal spermatozoon from a healthy control man (i). H&E staining of the SPEF2-mutated spermatozoa showed anomalies of the sperm flagella, including absent (ii), coiled (iii), short (iv) and irregular flagella (v). Scale bars: 5 µm. TEM analysis was conducted in the spermatozoa from a normal control man (vi) and the SPEF2-mutated probands (vii). The sperm defects observed in the SPEF2-mutated subject A028-II-1 are given as examples. Scale bars: 200 nm. CP, central pair of microtubules; IFT20, intraflagellar transport 20; M1, mutation 1; M2, mutation 2; M3, mutation 3; MMAF, multiple morphological abnormalities of the sperm flagella; MT, peripheral microtubule doublet; NK, nucleotide kinase; SMC, structural maintenance of chromosomes; SPEF2, sperm flagellar 2; TEM, transmission electron microscopy; WT, wild type.
Besides the Chinese cohort of MMAF, we also analysed the human exome data from an independent cohort with 78 subjects with MMAF of North African, French and Iranian origins.4 We identified another homozygous SPEF2 frameshift variant (c.3240delT, 1080Leufs*2) in an Iranian subject from a consanguineous family F003 (figure 1A). These three SPEF2 variants from MMAF-affected men were absent or extremely rare in the human genome data sets of more than 139 000 individuals from the 1000 Genomes Project, the Exome Aggregation Consortium and the Genome Aggregation Database (table 1). Furthermore, the mutation positions p.Arg304*, p.Ile1134Serfs*13 and p.Phe1080Leufs*2 in SPEF2 are highly conserved across species (online supplementary figure 1).
Homozygous stop-gain variants of SPEF2 identified in the men with MMAF
The long canonical transcript (T202 according to the GRCh38 version; online supplementary figure 2) of human SPEF2 gene encodes a sperm flagellar protein of 1822 amino acids, which is abundantly expressed in the testis.17 The SPEF2 protein contains a calponin homology (CH) domain at the N-terminus, a nucleoside/nucleotide kinase superfamily, a structural maintenance of chromosomes superfamily, an intraflagellar transport protein 20 (IFT20) binding domain, and an EF-hand motif in the C-terminus (figure 1B). Notably, all the three SPEF2 stop-gain variants identified in our MMAF cohorts affected the canonical transcript T202 (online supplementary figure 2). These variants were predicted to cause premature translational termination of SPEF2 and result in the truncation of the IFT20 binding domain, which is important for the interaction between SPEF2 and the IFT20 protein with a vital role in sperm tail development.18
Semen parameters of the MMAF cases carrying homozygous SPEF2 variants were investigated in this study. No spermatozoa with progressive motility were observed in any subjects affected by homozygous SPEF2 stop-gain variants (table 2). More than 65% of the immotile spermatozoa displayed abnormal flagella. Subject A028-II-1 with the early truncation mutation of SPEF2 c.910C>T (p.Arg304*) has a more severe malformation rate of sperm flagella than the other two subjects with late truncation mutations. Additionally, the sperm concentration and motility of these three SPEF2-mutated subjects were significantly lower than those of the DNAH1-mutated cases in this study (online supplementary table 2).
Semen characteristics and sperm morphology in the SPEF2-mutated men
H&E staining and DICM were used to assess sperm morphology. Obvious MMAF phenotypes were revealed in the spermatozoa from the SPEF2-mutated men when compared with the long and thin tails in the normal spermatozoa from a healthy control man (figure 1C, online supplementary figure 3). TEM was employed to investigate sperm flagellar ultrastructure of the SPEF2-mutated men. In the control specimen, sperm flagella showed the typical ‘9+2’ microtubule structure (nine peripheral microtubule doublets and a central pair of microtubules). However, all of the analysed cross sections of sperm flagella from SPEF2-mutated men were abnormal. The major morphological defect was the absence of the central pair complex (9+0 conformation) at the principal piece of sperm flagella (figure 1C).
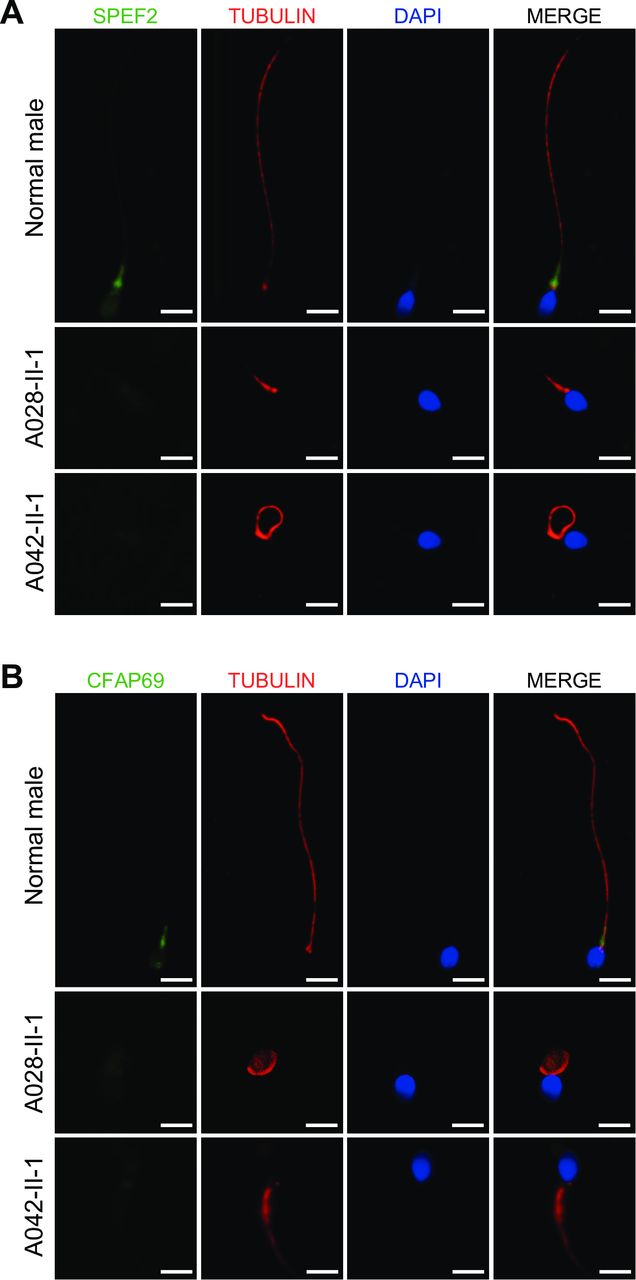
The pathogenicity of the MMAF-associated SPEF2 variants was further investigated at the protein level. We examined the distribution of SPEF2 and its related protein CFAP69 in the spermatozoa from normal and SPEF2-mutated men using immunofluorescence staining.5 In the spermatozoa from a normal control man, it was shown that SPEF2 localised to the base of sperm flagella. In contrast, SPEF2 immunostaining was almost absent in the spermatozoa from the homozygous SPEF2-mutated subjects (figure 2A). Furthermore, we also examined the localisation of CFAP69, a protein required for sperm flagellar assembly.5 In the normal control spermatozoa, CFAP69 immunostaining appeared strongly concentrated in the mid-piece of the sperm flagella (figure 2B), likely corresponding to mitochondrial sheath as reported by a recent study.5 In contrast, CFAP69 staining was very weak or almost absent in the SPEF2-mutated spermatozoa, while CFAP69 remained detectable in the sperm flagella from the cases with mutations in CFAP251 and DNAH1, respectively (figure 2B, online supplementary figure 4).

{kind=link}
{kind=link}
Localisation of SPEF2 and its related protein CFAP69 in the spermatozoa. (A) SPEF2 immunostaining in human spermatozoa from normal and SPEF2-mutated men. The spermatozoa were stained with anti-SPEF2 (green) against peptides at 227-275aa and anti-α-tubulin (red). DNA was counterstained with DAPI (4′,6-diamidine-2′-phenylindole dihydrochloride) as a nuclear marker. In the spermatozoa from a normal male, SPEF2 immunostaining is mainly concentrated at the basal body of the sperm flagellum. In contrast, SPEF2 staining is very weak or approximately absent in the sperm flagella from the SPEF2-mutated men. The α-tubulin staining uniformly displays the full-length flagellum in the normal male, whereas the sperm flagellar morphologies are abnormal in the SPEF2-mutated men. (B) Affected CFAP69 immunostaining in human spermatozoa from the SPEF2-mutated men. The spermatozoa were stained with anti-CFAP69 (green) and anti-α-tubulin (red) antibodies. In the spermatozoa from a normal male, CFAP69 staining appears to be mainly located in the mid-piece of the sperm flagellum. However, in the spermatozoa from the SPEF2-mutated men, CFAP69 staining is strongly reduced or absent. Abnormal flagellar morphologies could also be detected in the SPEF2-mutated spermatozoa. Scale bars: 5 µm. SPEF2, sperm flagellar 2.
Taken together, our genetic analyses and functional assays demonstrated the pathogenicity of homozygous SPEF2 stop-gain variants and suggested the involvement of SPEF2 in human MMAF.
Discussion
Spermatogenesis includes many testis-specific processes that are controlled by complex regulatory mechanisms.19 Normal organisation of the sperm tail is essential for sperm motility. In mice, SPEF2 staining was previously reported in the Sertoli cell cytoplasm, and seemed concentrated around the elongating spermatid heads just before spermatid bundle formation and at the stage with existing spermatid bundles.18 A recent study in mice also suggested that SPEF2 plays an important role in microtubule-mediated transport in the elongating spermatids to ensure proper male germ cell differentiation.17 Similarly, SPEF2 is involved in the development of cilia. The deficiency of SPEF2 in mice results in spermatogenic defects.20
Besides the molecular evidence in mouse models, an intronic insertion in the pig SPEF2 gene (also known as KPL2) resulted in aberrant splicing and led to immotile short-tail sperm defect in Finnish Yorkshire boars.21 Furthermore, a genetic variant c.2851G>T of SPEF2 in Holstein bulls induced aberrant splicing, which was involved in semen deformity and decreased sperm motility after cryopreservation.22 These experimental observations in animals are consistent with the human data of this study. The probands with homozygous stop-gain variants of SPEF2 showed sperm flagellar malformations and impaired motility (table 2), comparable with what had been described in the different animal models reported in previous studies. Our TEM results also revealed a complete absence of the central pair complex (9+0 conformation) at the principal piece of sperm flagella (figure 1C), suggesting an essential role for SPEF2 in normal axoneme formation.
Intriguingly, SPEF2 is likely to be functionally related to CFAP69, a protein that has been recently reported as an MMAF-associated factor.5 The levels of SPEF2 protein were described to be strongly reduced or absent in the spermatozoa from CFAP69-mutated men with MMAF.5 Consistently, here we observed a reduced and atypical staining of CFAP69 in the spermatozoa from SPEF2-mutated men, while CFAP69 staining is normal in the spermatozoa with mutations in other MMAF-associated genes such as CFAP251 and DNAH1. This evidence suggested that SPEF2 could be involved in sperm flagellar assembly together with CFAP69 via a specific but undetermined mechanism. Previous mass spectrometry analyses revealed potential interaction of SPEF2 with CFAP69 (Q8BH53), while the specific interaction mechanism is under investigation.17 A similar interaction can also be seen between cytoplasmic dynein 1 and SPEF2. SPEF2 could act as a linker protein for dynein 1-mediated cargo transport along the microtubules.17 Therefore, our experimental observations are in good agreement with previous evidence and confirm the important role of SPEF2 in normal sperm tail assembly.
Mutations at different positions of SPEF2 are likely to produce variable phenotypes. Male infertility and PCD-like symptoms were previously reported in a mouse model with Spef2 mutations.20 This previous study proposed that the PCD-like phenotypes could be caused by a missense mutation in exon 3 of mouse Spef2, while a stop-gain mutation at the C-terminus could be responsible for the spermatogenic defects.20 Similarly, ciliary abnormalities leading to bone growth defects and PCD-like symptoms were observed in the mice with a stop codon located after the Spef2 exon 2.23 Both of these PCD-associated Spef2 mutations in mice affected the N-terminus CH domain involved in regulating ciliary function (online supplementary figure 2).24 In contrast, a genetic variant (c.2851G>T) in SPEF2 exon 20 of Holstein bulls and the L1 retrotransposon insertion into SPEF2 intron 30 of Finnish Yorkshire boars only caused sperm tail defects.21 22 Therefore, PCD is not a common phenotype for SPEF2 mutations across species.
In our study, clinical examinations revealed no obvious PCD-like symptoms in men with SPEF2 truncation mutations. Among these cases, chest X-ray image and olfactory test with smelling bottles were further conducted on SPEF2-mutated subject A028-II-1. No obvious lung defects (such as bronchitis and pneumonia) or heart abnormalities were found (online supplementary figure 5).
Phenotypic variability observed among SPEF2-mutated animals/humans may be dependent on the distinct SPEF2 transcripts resulting in SPEF2 proteins with different functional domains. In humans, the alternative splicing of SPEF2 leads to multiple protein-coding transcripts (T201, T202, T203 and T212) with different expression levels between tissues (online supplementary figures 2 and 6). For example, the long canonical transcript T202 is preferentially expressed in sperm, suggesting an important role of the resulting protein in spermatogenesis. According to the data of the National Center for Biotechnology Information, Ensembl and a previous study,18 the proteins encoded by alternative SPEF2 transcripts contain different domains. The full-length SPEF2 protein encoded by transcript T202 contains several different domains, including the CH domain at the N-terminus and the IFT20 binding domain at the C-terminus (figure 1B). Additionally, the high similarity of amino acid sequences between the proteins encoded by long SPEF2 transcripts T202 and T203 suggests that the protein encoded by transcript T203 also includes the IFT20 binding domain (online supplementary figure 7). IFT20 has been reported to interact with SPEF2, and these two proteins colocalise in the Golgi complex and manchette of elongating spermatids, which play a vital role in sperm tail development.18 In this study, all three stop-gain variants of SPEF2 in human MMAF cases consistently affect the long transcripts T202 and T203, leading to the loss of IFT20 binding domain (online supplementary figure 2). Coincidentally, our RT-PCR assays detected the expression of these long SPEF2 transcripts in the sperm samples of healthy controls, while their expression was severely affected by the SPEF2 truncation mutation c.910C>T (p.Arg304*) (online supplementary figures 6 and 8). Therefore, it is speculated that deficiency of the full-length SPEF2 protein containing the IFT20 binding domain may potentially cause MMAF phenotypes by affecting the IFT20–SPEF2 interaction and intraflagellar transport, both of which are essential for flagella development and maintenance.25
The short SPEF2 transcript T212 was not affected by any of the three SPEF2 stop-gain variants in human MMAF of this study (online supplementary figure 2). This transcript is strongly expressed in human brain and lung (online supplementary figure 6). Remarkably, the short-length SPEF2 protein encoded by transcript T212 retained the CH domain that was previously described to have a critical role in regulating ciliary function.24 Intriguingly, in the mouse models with Spef2-associated PCD-like phenotypes, both pathogenic mutations affected all transcripts of mouse Spef2 and destroyed the CH domain at the N-terminus (online supplementary figure 2). The variants which do not alter the SPEF2 N-terminus, described here and in previous animal studies (Holstein bulls and pigs), consistently caused sperm tail defects without PCD symptoms. Therefore, the integrity of short SPEF2 transcripts (eg, T212 in humans) may result in a protein presenting a potentially functional SPEF2 N-terminus and avoid the manifestation of PCD phenotypes. Coincidentally, our RT-PCR assay demonstrated that this short SPEF2 transcript T212 remained normally expressed in the SPEF2-mutated subject A028-II-1 (online supplementary figure 8). Besides the different SPEF2 transcripts with variable functions, there may be other unknown mechanisms underlying the PCD phenotypes in the Spef2-mutated mice.
In conclusion, our genetic studies on human MMAF identified three homozygous stop-gain variants of SPEF2 encoding a sperm flagellar protein essential for spermatogenesis at the spermatid elongation stage. Our experimental observations, together with previously reported evidence in animals, strongly support the importance of SPEF2 in sperm motility and flagellar development. The differential expression of SPEF2 transcripts between tissues and the transcript-dependent organisation of functional domains may partially explain the phenotypic differences (MMAF with/without PCD) in SPEF2-mutated humans and animals. Our findings revealed a new human MMAF-associated gene, which will be informative in the molecular diagnosis and genetic counselling of infertile men with oligoasthenoteratozoospermia.26 27
Acknowledgments
We would like to thank the families for participating and supporting this study. We also thank the Center of Cryo-Electron Microscopy at Zhejiang University for technical support.
References
Footnotes
ChL, ML, XH, YoZ and PR contributed equally.
Contributors ChL, ML, XH, PR and FZ designed the study. ML, XH, AA-Y, HW, JZ, QT, Y-JZ, YaZ, ZZ and YC provided patients’ data and performed clinical assessments. ChL, YoZ, AA-Y, WeL, Z-EK, WaL and STi conducted the experiments. ChL, ML, XH, YoZ, HW, Z-EK, STa, CaL, LJ, PR, FZ and YC analysed the data. ChL, XH, PR, FZ and YC wrote the manuscript. PR, FZ and YC supervised the study.
Funding This work was supported by the National Natural Science Foundation of China (31625015, 31521003 and 81601340), Special Foundation for Development of Science and Technology of Anhui Province (2017070802D150 and YDZX20183400004194), Foundation of the Education Department of Anhui Province (KJ2016A370), Natural Science Foundation of Anhui Province (1708085QC59), Shanghai Medical Center of Key Programs for Female Reproductive Diseases (2017ZZ01016), and Shanghai Municipal Science and Technology Major Project (2017SHZDZX01).
Competing interests None declared.
Patient consent for publication All patients gave their signed informed consent for this study.
Ethics approval Declaration of Helsinki and approved by the Ethical Committees of the centers participating in this study.
Provenance and peer review Not commissioned; externally peer reviewed.