Article Text

Abstract
Background Male infertility is a major issue of human reproduction health. Asthenoteratospermia can impair sperm motility and cause male infertility. Asthenoteratospermia with multiple morphological abnormalities of the flagella (MMAF) presents abnormal spermatozoa with absent, bent, coiled, short and/or irregular-calibre flagella. Previous studies on MMAF reported that genetic defects in cilia-related genes (eg, AKAP4, DNAH1, CFAP43, CFAP44 and CFAP69) are the major cause of MMAF. However, the known MMAF-associated genes are only responsible for approximately 30% to 50% of human cases. We further investigated the cases with MMAF in search of additional genes mutated in this condition.
Methods and results We conducted whole exome sequencing in a male individual with MMAF from a consanguineous Han Chinese family. Sanger sequencing was also conducted in additional individuals with MMAF. Intriguingly, a homozygous frameshift mutation (p.Leu357Hisfs*11) was identified in the gene encoding CFAP69 (cilia and flagella-associated protein 69), which is highly expressed in testis. The subsequent Sanger sequencing of the CFAP69 coding regions among 34 additional individuals with MMAF revealed a case with homozygous nonsense mutation (p.Trp216*) of CFAP69. Both of these CFAP69 loss-of-function mutations were not present in the human population genome data archived in the 1000 Genomes Project and ExAC databases, nor in 875 individuals of two Han Chinese control populations. Furthermore, we generated the knockout model in mouse orthologue Cfap69 using the CRISPR-Cas9 technology. Remarkably, male Cfap69-knockout mice manifested with MMAF phenotypes.
Conclusion Our experimental findings elucidate that homozygous loss-of-function mutations in CFAP69 can lead to asthenoteratospermia with MMAF in humans and mice.
- CFAP69
- flagella
- knockout mice
- sequencing
- male infertility
Statistics from Altmetric.com
Introduction
Human infertility with multiple morphological abnormalities of the flagella (MMAF), the term of which was first named by Pierre F. Ray in 2014, mainly presents a combination of sperm flagellar malformations, including absent, bent, coiled, short and/or irregular-calibre flagella.1 2 In addition to these sperm morphological abnormalities that can be detected by light microscopy, lack of central microtubules and/or dynein arms were also observed by transmission electron microscopy (TEM) in the sperm flagella affected by MMAF.1 3 This kind of human asthenoteratospermia was previously reported as dysplasia of fibrous sheath (DFS) and short/stump tails. The incidence of MMAF or DFS has not been investigated precisely; however, it was estimated to be higher than one per 10 000–20 000 births of primary ciliary dyskinesia (PCD (MIM: 244400)), another disorder of motile cilia.4
Previous genetic studies on flagellar malformations have reported that functional defects in the cilia-related genes are the major causes of MMAF in humans and model organisms.2 5 However, only a few genes have been formally identified to cause human MMAF: AKAP4 (MIM: 300185), DNAH1 (MIM: 603332), CFAP43 (MIM: 617558), CFAP44 (MIM: 617559) and CFAP69 (MIM: 617949) (online supplementary table S1).1 2 5–7 According to the latest report,8 the known genetic defects could explain 31% of MMAF-affected individuals. Therefore, we conducted further genetic analyses to investigate the human individuals with MMAF in search of additional genes mutated in this condition.
Supplemental material
In this study, we analysed an MMAF case using whole exome sequencing (WES) and identified a homozygous mutation in CFAP69. This gene encodes the protein of cilia and flagella-associated protein 69, which contains WD repeat domains. Subsequently with Sanger sequencing of other 34 men with MMAF, we found another proband with homozygous CFAP69 mutation. The total contribution of CFAP69 accounts for 5.7% (2/35) of the MMAF subjects recruited in this study. Most importantly, we generated a Cfap69-knockout (KO) mouse model, which consistently presented with the MMAF phenotypes. Overall, our study further confirmed that homozygous loss-of-function mutations of CFAP69 are associated with MMAF and primary male infertility.
Materials and methods
Subjects
In this study, we enrolled a Han Chinese man with MMAF from a consanguineous family (A001 in figure 1) at the First Affiliated Hospital of Anhui Medical University for genetic analysis. Thirty-four additional Han Chinese individuals with MMAF were also investigated by Sanger sequencing. Informed consents were obtained from each subject.

Identification of homozygous CFAP69 mutations in the human individuals with multiple morphological abnormalities of the flagella (MMAF). (A) Two consanguineous families (A001 and A006) affected by MMAF associated with CFAP69 mutations. Homozygous loss-of-function mutations of CFAP69 were identified in the probands. Their parents carried the heterozygous mutations. Sanger sequencing plots were also shown for the probands and their parents. The mutant sequences were highlighted by yellow. WT, wild type. (B) The mutation positions in CFAP69 were also shown, which are conserved across species.
The proband (IV-1) in family A001 suffered from primary infertility for 5 years due to severe teratozoospermia with no obvious PCD-related symptoms such as sinusitis, bronchitis, pneumonia and otitis media. Semen volume, sperm concentration and motility were evaluated with Sperm Class Analyzer CASA System (Spain) according to the WHO guidelines (2010). Semen samples were stained by a modified Papanicolaou method for sperm morphological analysis. The percentage of morphologically normal and abnormal spermatozoa was evaluated according to the WHO guidelines. The morphological abnormalities of sperm flagella were classified into five categories as absent, short, bent, coiled flagella and flagella of irregular calibre.1 One spermatozoon was classified to only one morphological category according to its major flagellar abnormality.3 7
Whole exome sequencing
Genomic DNA was extracted from peripheral blood sample using the DNeasy Blood and Tissue kit (QIAGEN). Agilent SureSelectXT Human All Exon Kit and Illumina HiSeq X-TEN platform were employed to sequence the human exome. The follow-up bioinformatic analysis was based on standard assembly (Burrows-Wheeler Aligner), calling, analysis (Genome Analysis Toolkit) and annotation (ANNOVAR).9–11
TEM analysis
TEM was used to investigate the sperm flagellar ultrastructures in the human individuals affected by MMAF. The sperm cells were fixed routinely. After embedded in Epon 812, ultrathin sections were stained with uranyl acetate and lead citrate, and photographed under TEM (TECNAI-10; Philips) with an accelerating voltage of 80 kV.
KO mouse model
The mouse model was generated using the CRISPR-Cas9 technology based on the C57BL/6J mouse strains.12 The protocol of genome editing was provided by Nanjing Biomedical Research Institute of Nanjing University. These sgRNAs were phosphorylated and ligated to the pUC57-gDNA-T7 plasmid that had been digested with BsaI already. Single colonies were picked out from each transformation plate and identified by PCR and sequencing. After in vitro transcription and purification, the plasmid was injected into the cytoplasm of fertilised eggs, which were generated by superovulated female C57BL/6J mice mated with wild-type (WT) C57BL/6J males. Blastocysts were transferred into pseudo-pregnant female mice. This study was carried out in accordance with the recommendation in the Guide for the Care and Use of Laboratory Animals of the National Institutes of Health.
Histological analysis of mouse tissues
Fresh mouse testis and epididymis were fixed in modified Davidson’s fluid (MDF: 50% diluted water, 30% formaldehyde, 15% ethanol, 5% glacial acetic acid) for over 48 hours, embedded in paraffin. Sections were cut at a 3 µm thickness, then deparaffinised in 65°C overnight. After deparaffinisation, slides were stained with H&E, dehydrated and mounted.
Immunofluorescence analysis
Both human spermatozoa and mouse testis sections were stained by immunofluorescent. Human spermatozoa and mouse testis tissues were fixed in 4% paraformaldehyde and Bouin’s solution, respectively. Human spermatozoa were coated on the slides and incubated with CFAP69 polyclonal antibody (1:100, bs-15278R-A647, Bioss) and acetylate tubulin (1:1000, T6793, Sigma) at 4°C overnight. AlexaFluor 647 anti-Rabbit (1:1000, 34213ES60, Yeasen) and AlexaFluor 488 anti-Mouse (A21206, Invitrogen) were applied for an hour incubation at room temperature. Mouse testis tissues were embedded in paraffin and sectioned. After being carefully deparaffinised, the tissue sections were put into 10 mM citrate buffer (pH 6.0) and boiled for 10 min. Then the slides were incubated with anti-acetylated-α-tubulin (1:1000, T6793, Sigma) at 4°C overnight, and with AlexaFluor 488 anti-Mouse (A21206) for additional 2 hours at room temperature. Finally, both human spermatozoa and mouse testis tissues slides were coated with Hoechst 33342 for 5 min. After mounted with VectaShield, slides were observed with an LSM700 confocal microscope (Carl Zeiss).
Results
The semen analysis in the proband (IV-1) of family A001 demonstrated that the sperm concentration is normal, but the rate of spermatozoa with progressive motility is only 0.8% (table 1). The morphological analysis showed typical MMAF phenotypes as follows: absent flagella (13%), short flagella (28%), coiled flagella (35%), angulation (7.5%) and flagella of irregular calibre (2.5%) (table 1 and figure 2). WES was conducted for genetic analysis. Due to the consanguineous status of family A001, we assumed that the causative mutation could be a rare homozygous mutation in human populations.13 14 Loss-of-function variants and potentially deleterious missense variants predicted by SIFT, PolyPhen-2 and MutationTaster were prioritised.15–17 In addition, the genes with significant expression in testis and potential roles in spermatogenesis were preferred. Intriguingly, we identified a homozygous frameshift mutation c.1069_1070insAC (p.Leu357Hisfs11*) of CFAP69 (alternatively termed C7orf63, NM_001039706.2) in the proband (IV-1) of family A001 (figure 1). This mutation was verified by Sanger sequencing, and both parents are carriers of the heterozygous mutation (figure 1). This CFAP69 frameshift mutation was absent from the human population genome data of more than 62 000 individuals archived in the 1000 Genomes Project and ExAC databases (online supplementary table S2).13 14 We also investigated two control populations with 300 and 575 individuals of Han Chinese ancestry, respectively. Notably, the CFAP69 mutation c.1069_1070insAC is also absent from the ethnically matched control populations (online supplementary table S2). These findings suggested that homozygous loss-of-function mutation of CFAP69 could be pathogenic.

Light microscopy showed morphological abnormalities of the human spermatozoa affected by homozygous CFAP69 mutations. (A) The light microscopy image for a spermatozoon with normal morphology from a healthy man. (B and C) The human spermatozoa affected by homozygous CFAP69 mutation presented multiple morphological abnormalities of the flagella phenotypes, such as short and coiled flagella.
Semen characteristics and sperm morphology in the human individuals with homozygous CFAP69 mutations and those with unknown causes
To further investigate the potential contribution of other CFAP69 loss-of-function mutation(s) to human MMAF, we sequenced the coding regions of CFAP69 using Sanger sequencing in 34 additional Han Chinese individuals with MMAF. Nine of them were enrolled from consanguineous families. These male individuals were all affected by primary infertility due to multiple flagella malformations (absent, bent, coiled, short flagella or irregular calibre of flagella). The rates of abnormal flagella range from 77.5% to 100% among these 34 individuals with MMAF. The symptoms of PCD, including sinusitis, bronchitis, pneumonia and otitis media, were carefully excluded. Intriguingly, we identified another homozygous loss-of-function mutation of CFAP69 (c.647G>A, p.Trp216*) in the proband (IV-1) of family A006 (figure 1). The major MMAF phenotypes in this case are short and coiled flagella (47.5% and 32.5% among spermatozoa, respectively) (table 1). Sanger sequencing confirmed that this homozygous nonsense mutation of CFAP69 was parentally inherited from heterozygous carriers (figure 1). We also confirmed that this CFAP69 nonsense mutation was absent from the available human population data, including those from the public databases of the 1000 Genomes Project and ExAC,13 and our in-house data of two Han Chinese control populations (online supplementary table S2). Therefore, CFAP69 homozygous loss-of-function mutations account for 5.7% (2/35) of the MMAF subjects included in this study.
To investigate the roles of CFAP69 in sperm flagellar formation, we examined the location of CFAP69 protein in sperm flagella from control and CFAP69-deficient men by immunofluorescence. It was shown that the CFAP69 protein was located in the midpiece of sperm flagella of a normal man (as a positive control), whereas the CFAP69 signal was absent from the sperm flagella of the CFAP69-deficient men (figure 3). Furthermore, we used TEM to examine the sperm flagellar ultrastructures in the CFAP69-deficient men. To assess microtubule assembly, over 50 flagellar cross-sections were investigated. TEM showed the typical ‘9+2’ microtubule structure (nine peripheral microtubule doublets paired with nine outer dense fibres and the central pair of microtubules) in normal spermatozoa from a healthy man (figure 4A). However, the central pair of microtubules were absent in most spermatozoa from the men affected by CFAP69 homozygous loss-of-function mutations (figure 4B,C).
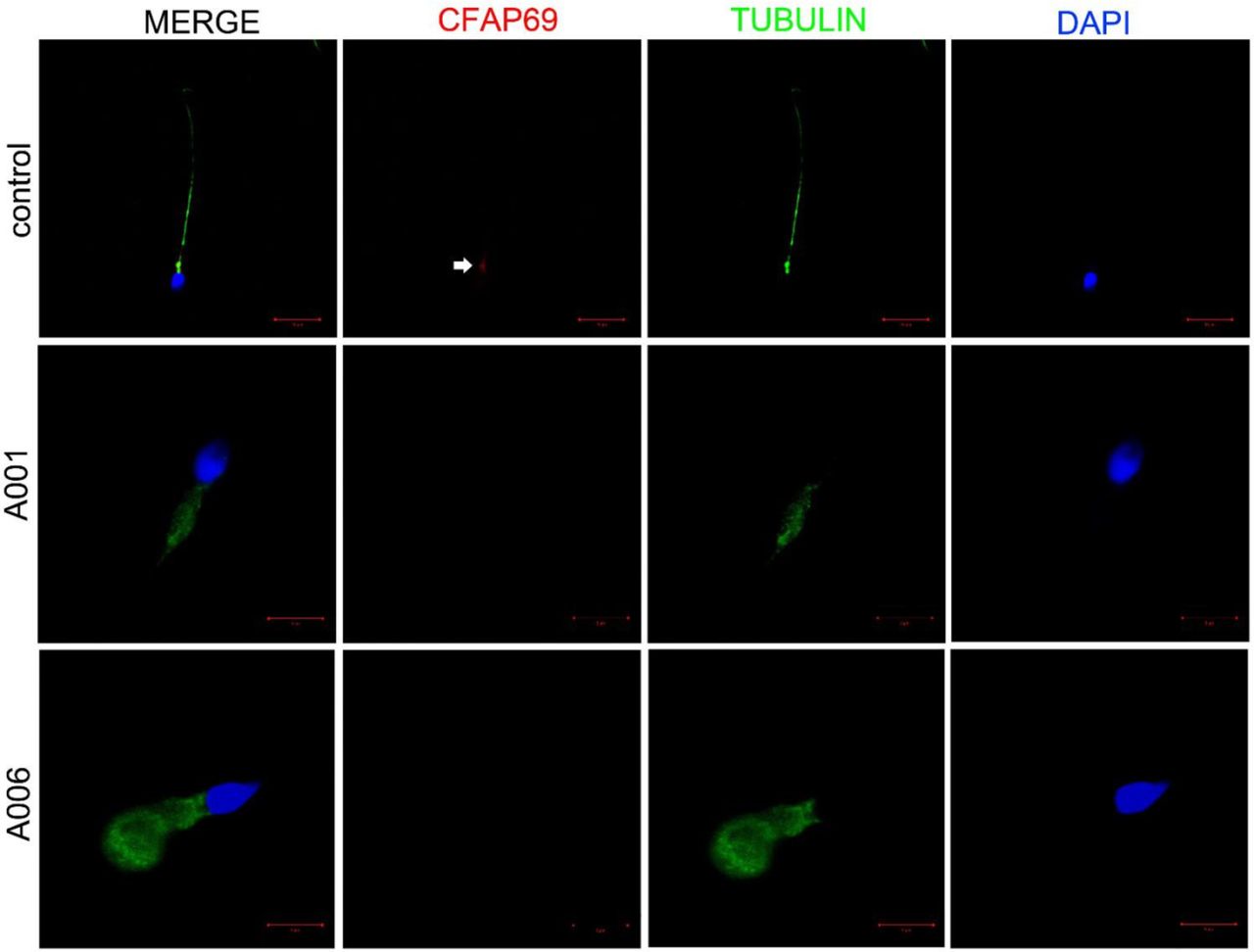
Immunofluorescence staining in CFAP69-deficient men reveals an absent CFAP69 protein. Spermatozoa from a fertile control and two CFAP69-deficient men are stained with anti-CFAP69 (red) and anti-acetylated tubulin (green) antibodies. DNA was counterstained with DAPI (4′,6-diamidino-2-phenylindole). In the fertile control, the CFAP69 immunostaining (red) is concentrated in the midpiece of the sperm (white arrow) while the CFAP69 signal is absent in sperm flagella from both CFAP69-deficient men. Scale bars: 10 µm and 5 µm.
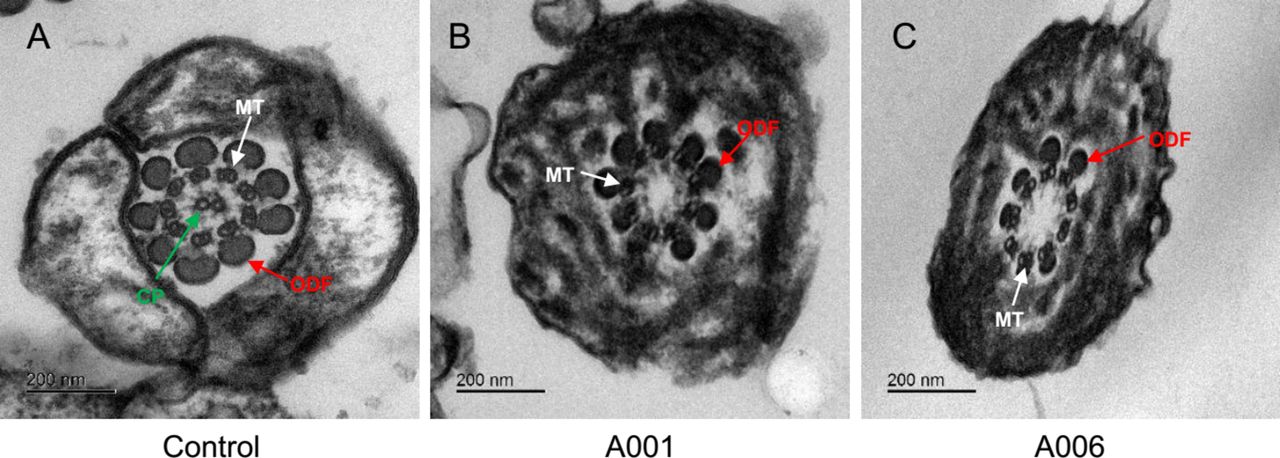
Abnormal human sperm ultrastructures caused by homozygous CFAP69 mutations. (A) The ultrastructure of cross-section in a normal spermatozoon from a healthy man. Transmission electron microscopy showed the typical ‘9+2’ microtubule structure: nine peripheral microtubule doublets paired with nine outer dense fibres and the central pair of microtubules. (B and C) The absence of central pair of microtubules was detected in most spermatozoa affected by homozygous CFAP69 mutation. The cross-sections showed abnormal spermatozoa with ‘9+0’, normal arrangement of peripheral microtubules and lack of the central pair of microtubules. CP, the central pair of microtubules (green arrows); MT, peripheral microtubule doublet (white arrows); ODF, outer dense fibre (red arrows). Scale bars: 200 nm.
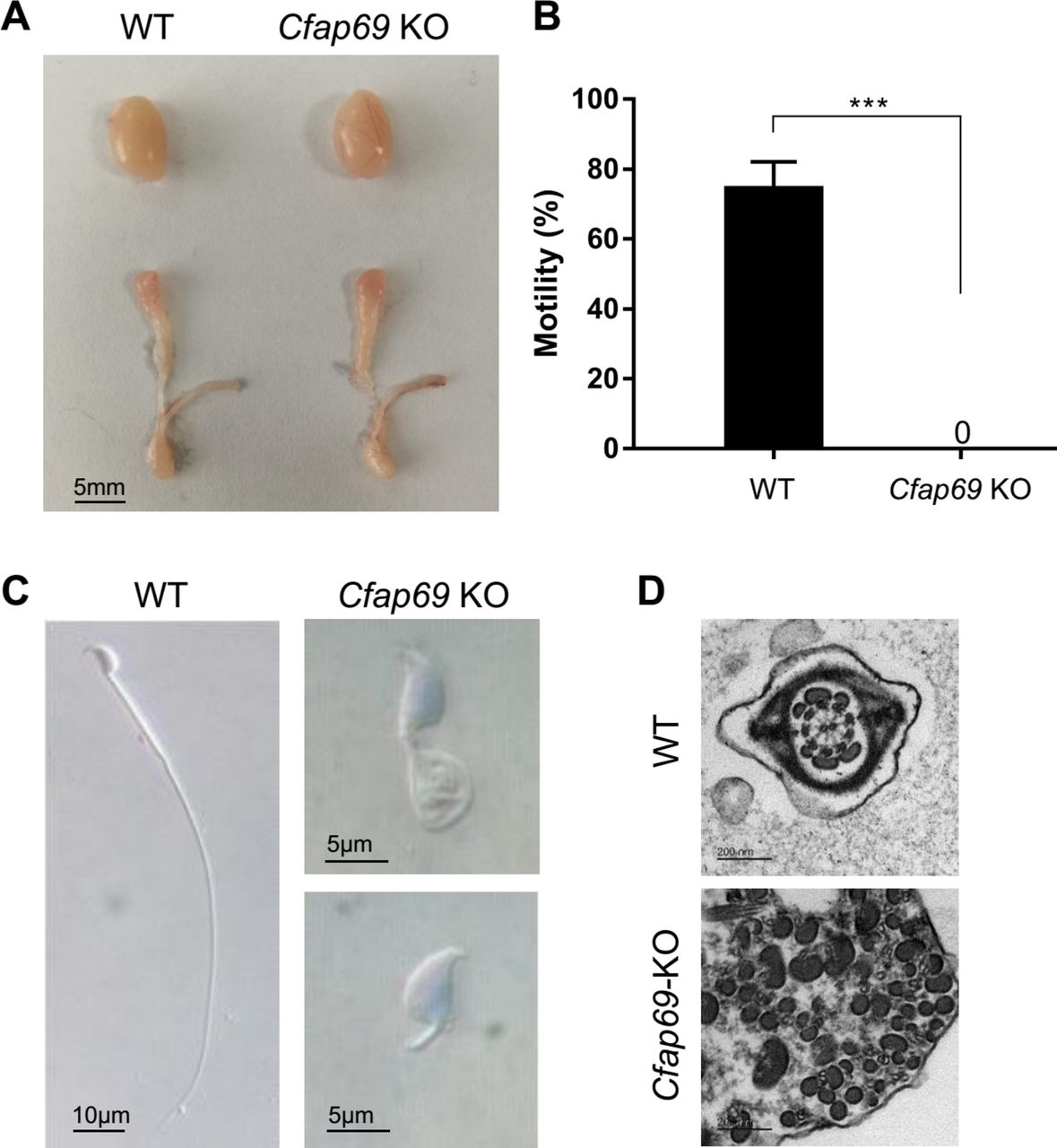
To better understand the roles of mouse orthologue Cfap69 in sperm flagellar formation, the Cfap69-KO mice were generated using the CRISPR-Cas9 technology. The sgRNAs were designed on CRISPR Design (online supplementary table S3). Three Cfap69 exons (2 to 4) were deleted in the mutated mice (online supplementary figure S1). This Cfap69 exonic deletion resulted in a frameshift mutation, which was confirmed by Sanger sequencing of the mutated cDNA. Fertility was tested in the male mice of the different genotypes (8–12 weeks, n=6). Each male mouse was caged with two WT C57BL/6J females (8–12 weeks), and vaginal plug was checked every morning. Once a vaginal plug was identified (day 1 postcoitus), the male was allowed to rest for 2 days, after which another female was placed in the cage for another round of mating. The plugged female was separated and single caged, and the pregnancy results were recorded. If a female did not generate any pups by day 22 postcoitus, it was deemed as not pregnant and euthanised to confirm that result. The fertility test lasted for 3 weeks. The average litter size of the WT mice was 9.65±0.21 and the Cfap69-KO mice had no offspring.
To clarify the reason of infertility, semen analysis, morphological study, H&E staining and immunofluorescent staining of testis have been conducted in detail. The gross evaluation of testis and epididymis did not show any obvious differences between male Cfap69-KO and WT control mice (figure 5A). Computer-assisted sperm analyzer (CASA, Cyto-S, VideoTesT) was used to assess mouse semen samples collected from cauda epididymis. Semen samples from each epididymis were diluted in 1 mL sperm rinse (10101, Vitrolife) and examined after incubation for 30 min at 37°C. CASA analysis showed the dramatic decrease of the motility of Cfap69-KO mice, revealing total immotility which was significantly distinctive from WT mice (figure 5B, online supplementary table S4). We also investigated the Papanicolaou-stained spermatozoa under light microscopy. Remarkably, no normal sperm flagella were observed for male Cfap69-KO mice. Their typical morphological abnormalities of sperm flagella included absent, short, coiled, bent and irregular-calibre flagella (figure 5C). Moreover, we also observed that the morphological abnormality rate of sperm head in the Cfap69-KO mice was significantly higher than that of the WT mice. The rates of abnormal sperm head were up to 89.5%; the main abnormal types were abnormal head shape and acrosome abnormality. In addition, we also performed the TEM in the spermatozoa from the Cfap69-KO mice. Severer than the humans, the microtubule doublets and outer dense fibres were scattered with chaos (figure 5D). The disordered pattern indicated that mouse Cfap69 also played vital roles in the structure arrangement of sperm flagella.

Cfap69 knockout (KO) leads to male infertility and sperm flagellar malformation in mice. (A) No obvious difference in the sizes of the testis and epididymis was found between the wild-type (WT) and Cfap69-KO mice. (B) The histogram displayed the distinctive sperm motility between WT and Cfap69-KO mice. The motility of male Cfap69-KO mice declined apparently to zero, suggesting the complete motility defects. (C) The light microscopy image of a spermatozoon with normal elongated flagellum from a male WT mouse. No normal sperm flagella were observed in male Cfap69-KO mice. The sperm flagellar abnormalities shown by light microscopy were coiled, short flagella and other distorted shapes in accordance with the clinical phenotypes in the human individuals with multiple morphological abnormalities of the flagella. (D) Transmission electron microscopic analyses of WT and Cfap69-KO spermatozoa. Compared with WT, the microtubule doublets and outer dense fibres were scattered with chaos in Cfap69-KO spermatozoa. ***P<0.001, Scale bars: 5 mm, 10 µm, 5 µm and 200 nm.
To further investigate the flagella defects in the Cfap69-KO mice, testis sections were stained with acetylate tubulin by immunofluorescent. The acetylated tubulin signal can be clearly observed in the seminiferous tubules from the WT mice (figure 6A,C), while relatively few signal could be detected in the Cfap69-KO mice (figure 6B,D). H&E staining of testis was also performed to assess spermatogenesis of Cfap69-KO and WT mice. No obvious differences of seminiferous tubule structure and spermatogenic cells were observed between male Cfap69-KO and WT mice (online supplementary figure S2). However, the testicular and the epididymal spermatozoa of the Cfap69-KO mice presented short flagella (online supplementary figure S2B,D), while the spermatozoa from the male WT mice had normal elongated flagella (online supplementary figure S2A,C).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Cfap69 knockout (KO) mice showed sperm flagellar malformation. Immunofluorescence analysis of the localisation of acetylated tubulin in elongated spermatids from the seminiferous tubules at stages VII–VIII. (A, C and E) Wild-type (WT) mice. (B, D and F) Cfap69-KO mice. Complete axoneme signals can be observed in the testes of the WT mice, while relatively few abnormal signals could be observed in testes of the Cfap69-KO mice, indicating the defected flagellar formation. DAPI, 4′,6-diamidino-2-phenylindole.
Discussion
Sperm flagella are assembled during the last step of spermatogenesis called spermiogenesis. Flagellar proteins typically are synthesised during spermiogenesis and then must be transported and assembled into the developing flagellum, including MMAF-associated genes (online supplementary table S1).18 Principal piece localisation and immunogold signal of AKAP4 showed it is necessary to correctly assemble into longitudinal columns and transverse ribs of the fibrous sheath in human. All of DNAH1, CFAP43 and CFAP44 proteins expressed throughout flagellum, suggesting that they are the essential components of axoneme. Although lack of immunoelectron microscopy analysis, DNAH1 was proposed to link inner dynein arms to radial spoke 3 through the defects of Dnah1-KO mice.1 CFAP43 and CFAP44 were also proved to be closely associated with the part of axoneme and most probably corresponding doublet of microtubules in Trypanosoma brucei model.19
CFAP69 is highly expressed in human testis according to the RNA-seq data of the ENCODE project and other gene expression data archived in the database of Expression Atlas.20 21 Furthermore, the CFAP69 protein was also identified as a testis-enriched protein using an in-depth proteomics analysis in human spermatozoa.22 The orthologue of CFAP69 in Chlamydomonas reinhardtii is FAP69 (flagellar protein 69), a 135 kDa protein containing two armadillo repeat domains.23 24 The experimental evidence from C. reinhardtii suggested that FAP69 is involved in the C1b projection of central pair microtubules in Chlamydomonas flagella. Taken together, these previous observations suggest that CFAP69 could play an important role in sperm flagellar formation.
Very recently, Dong et al. 8 reported a nonsense mutation and a splice-site mutation of CFAP69 in a North African and an Iranian case with MMAF, respectively. In our study, both of the pathogenic mutants in CFAP69 were loss-of-function variants (one stop-gain mutation and one frameshift mutation), which had significant damaging effects on CFAP69.
All of MMAF studies present typical MMAF manifestation, such as immotility and five major types of flagellar malformation in both known and unknown cases. Our data also showed no particular differences between CFAP69-deficient men and other cases with unknown genetic causes. However, the very low sperm concentration in CFAP69-deficient men is reported by Dong et al.8 This phenomenon may reflect the phenotypic variance between cases. For example, in the previous study of DNAH1-associated MMAF,1 three brothers (P1–P3) in one family had the same homozygous splicing mutation of DNAH1 but they manifested variable sperm concentrations. The sperm concentration is normal in P1, whereas the sperm concentrations were less than 5.0×106/mL for P2 and P3. It has been known that the factors affecting sperm concentrations are complicated, including environmental factors, endocrine factors, infection and so on. We speculated that the discrepancy of sperm concentration could be caused by these factors.
Despite the main defects in sperm flagella, many studies also showed the malformation of sperm head.8 We assessed the morphology of sperm head in the CFAP69-deficient men and Cfap69-KO mice by light microscopy. In accordance with the previous report, most of the individuals presented with obvious head morphological abnormalities. The abnormal rate of head sperm in CFAP69-deficient men were 95% and 97%, respectively (online supplementary table S5), and small acrosome or tapered heads were the main abnormal types. In addition, we also observed that the head morphological abnormality rate of Cfap69-KO mice was significantly higher than that of the WT mice. The malformation of sperm head and flagella in mice was particularly consistent with the sperm phenotypes of the CFAP69-deficient men with MMAF (figure 2 and table 1). Details of spermatid head formation mechanism are not fully explored, but parts of puzzle are starting to be speculated. The manchette, a microtubule-enriched perinuclear plays an essential role of assembling functional spermatozoa.25 It not only provides the physical forces for shaping sperm head but also regulates the transportation and processes of sperm flagellar assembly. The impaired component of transport or developing flagellar may also influence the shaping of sperm head. More patients and animal models with mutations in MMAF-associated genes should be investigated for the association with head shaping and flagellar developing.
PCD is a motile cilia disorder often to be linked to MMAF due to the similar conserved axoneme structure. The MMAF cases enrolled in this study were strictly excluded from no obvious PCD-related symptoms such as sinusitis, bronchitis, pneumonia and otitis media. The A001 proband conducted olfactory test with smelling bottles and chest CT scanning, but no obvious olfactory and lung defect was found (online supplementary figure S3). We also investigated the airway cilia by staining the bronchial tube cross-section of Cfap69-KO mice and found that the cilia and ciliated cells can be observed remarkably in both WT and Cfap69-KO mice (online supplementary figure S4). Although the cilia and flagella share lots of components and functions, it is possible that CFAP69 play different roles during flagella and cilia formation. It has been reported the axoneme composition is different, especially the specific periaxonemal structure, fibre sheath and outer dense fibres (ODFs) in flagella.26 Interestingly, CFAP69 is located in the midpiece of flagella where it is surrounded by mitochondrial sheath. Further studies should explore the different components and organisation of motile cilia and flagella.
In conclusion, our experimental findings in both human MMAF associated with CFAP69 homozygous loss-of-function mutations and male Cfap69-KO mice strongly suggest that CFAP69 plays a vital role in sperm flagellar formation. Genetic defects of CFAP69 can cause MMAF phenotypes and lead to male infertility in humans and mice. Homozygous pathogenic mutations of CFAP69 are very rare or absent from human control populations, but can be enriched in the male individuals affected by MMAF. Further investigations on MMAF-associated CFAP69 mutations may shed light on the molecular mechanism of sperm flagellar abnormalities and other ciliopathies.
Acknowledgments
We would like to sincerely thank all the patients with asthenoteratospermia and their families for their support and participation. We also thank the staff from the Model Animal Research Center of Nanjing University for supporting knockout mouse generation.
References
Footnotes
XH, WL and HW contributed equally.
Contributors XH: study design, molecular experiment and original draft preparation; WLi: data analysis, molecular experiment and original draft preparation; HWu: data collection, mouse knockout and original draft preparation; ML: molecular experiment; WLiu: molecular experiment and data analysis; CLiu: molecular experiment; FuZ: data analysis; CLi: data analysis; YF: molecular experiment; CY: data analysis; HC: mouse knockout and molecular experiment; JZ: mouse knockout and molecular experiment; JT: sperm analysis; TC: molecular experiment; DT: data collection; BS: data collection; XW: data collection; XZ: data collection and sperm analysis; HWa: data analysis; ZW: data analysis; SY: data analysis; HS: molecular experiment and data analysis; PZ: data analysis; LJ: data analysis; JW: data analysis; ZZ: study conception and design; FeZ: study design, data analysis and editing of the manuscript; YC: study conception and design.
Funding This study was supported by Foundation of the Department of Science and Technology of Anhui Province (2017070802D150), National Key Research and Development Program of China (2016YFC0905100 and 2017YFC1001100), National Natural Science Foundation of China (31625015, 11627803, 81601340, 31521003, 31571297 and 31771396), Shanghai Medical Center of Key Programs for Female Reproductive Diseases (2017ZZ01016), Foundation of the Education Department of Anhui Province (KJ2016A370), Maternal and Child Health Research Project of Jiangsu Province (F201521) and Shanghai Municipal Science and Technology Major Project (2017SHZDZX01).
Competing interests None declared.
Patient consent Obtained.
Ethics approval Ethics committee of Anhui Medical University and the Institutional review boards of the School of Life Sciences at Fudan University.
Provenance and peer review Not commissioned; externally peer reviewed.