Abnormal Response of the Proliferation and Differentiation of Growth Plate Chondrocytes to Melatonin in Adolescent Idiopathic Scoliosis

{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
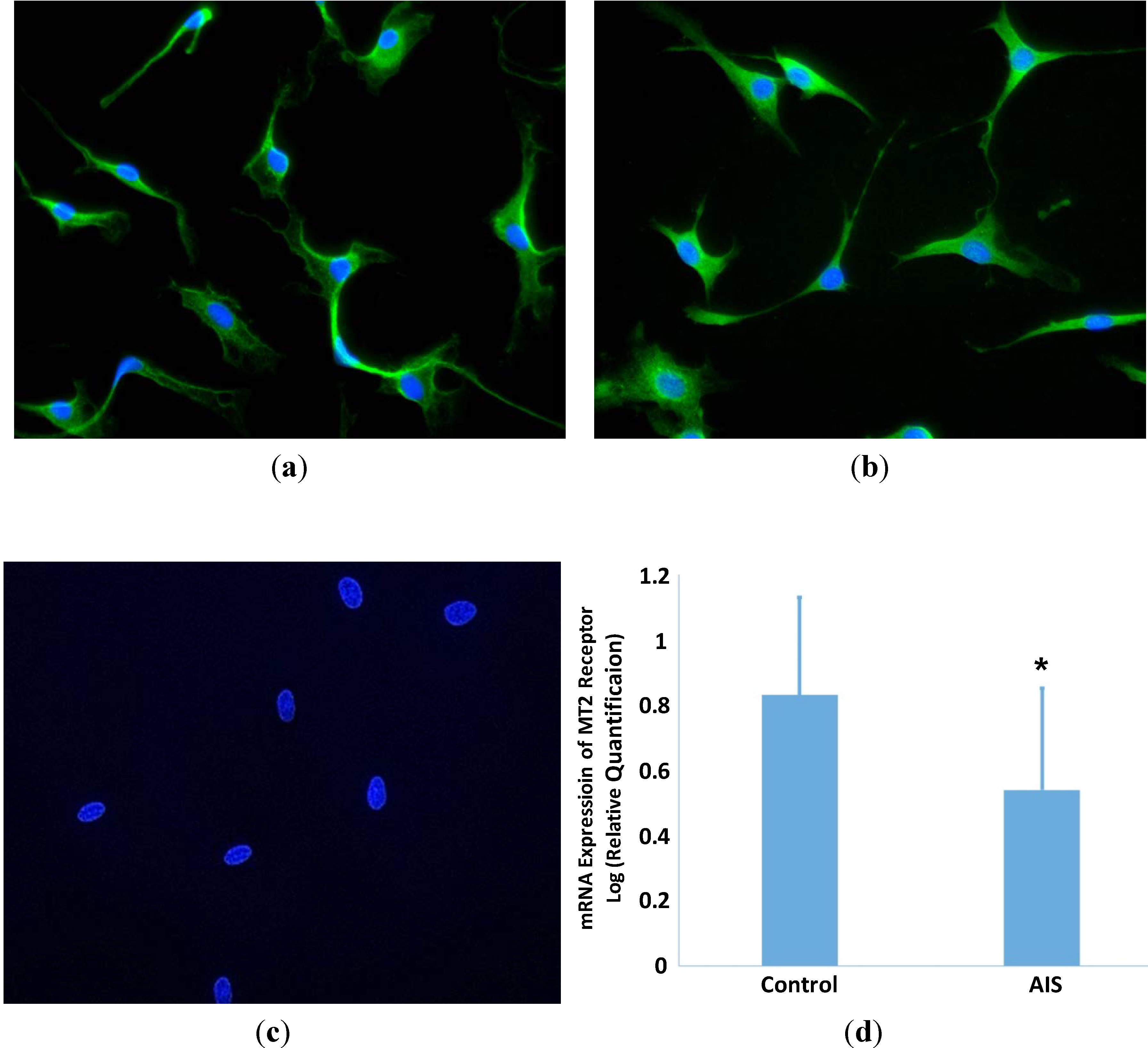
2.1. Expression of MT2 Receptor in GPCs

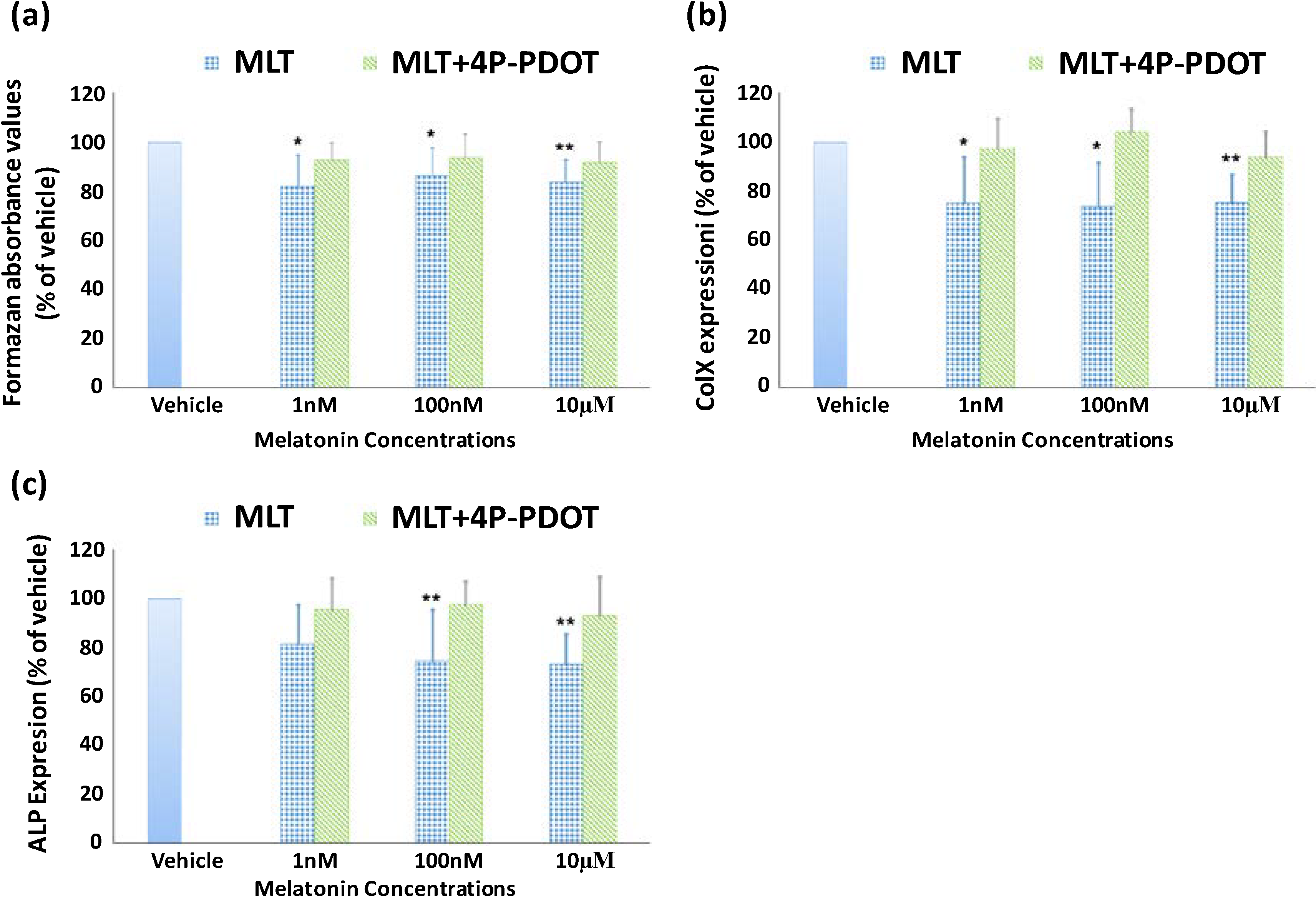
2.2. Effect of Melatonin on the Proliferation and Differentiation of Growth Plate Chondrocytes (GPCs) from Normal Control Subjects

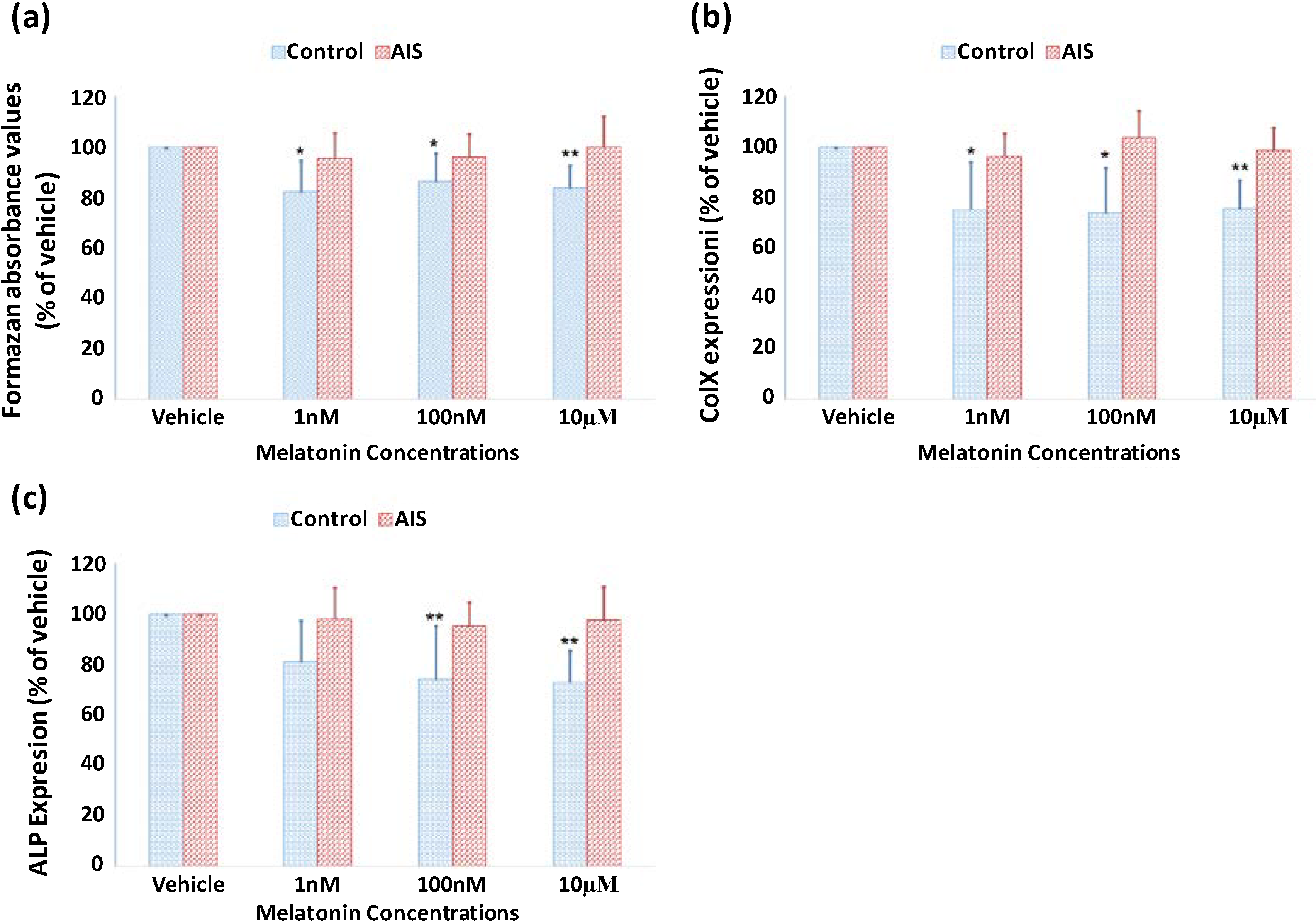
2.3. Lack of Response of GPCs to Melatonin Treatment in Both Proliferation and Differentiation in AIS Patients

3. Experimental Section
3.1. Reagents
3.2. Recruitment of Subjects
3.3. Primary Culture of GPCs
3.4. Expression of Melatonin Membrane MT2 Receptor
3.5. Effect of Melatonin on Proliferation of GPCs
3.6. Effect of Melatonin on Differentiation of GPCs
3.7. Statistical Analysis
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Weinstein, S.L.; Dolan, L.A.; Cheng, J.C.; Danielsson, A.; Morcuende, J.A. Adolescent idiopathic scoliosis. Lancet 2008, 371, 1527–1537. [Google Scholar]
- Burwell, R.G.; Dangerfield, P.H.; Freeman, B.J. Concepts on the pathogenesis of adolescent idiopathic scoliosis. Bone growth and mass, vertebral column, spinal cord, brain, skull, extra-spinal left-right skeletal length asymmetries, disproportions and molecular pathogenesis. Stud. Health Technol. Inform. 2008, 135, 3–52. [Google Scholar]
- Wang, W.J.; Yeung, H.Y.; Chu, W.C.; Tang, N.L.; Lee, K.M.; Qiu, Y.; Burwell, R.G.; Cheng, J.C. Top theories for the etiopathogenesis of adolescent idiopathic scoliosis. J. Pediatr. Orthop. 2011, 31, S14–S27. [Google Scholar]
- Siu King Cheung, C.; Tak Keung Lee, W.; Kit Tse, Y.; Ping Tang, S.; Man Lee, K.; Guo, X.; Qin, L.; Chun Yiu Cheng, J. Abnormal peri-pubertal anthropometric measurements and growth pattern in adolescent idiopathic scoliosis: A study of 598 patients. Spine 2003, 28, 2152–2157. [Google Scholar]
- Wei-Jun, W.; Xu, S.; Zhi-Wei, W.; Xu-Sheng, Q.; Zhen, L.; Yong, Q. Abnormal anthropometric measurements and growth pattern in male adolescent idiopathic scoliosis. Eur. Spine J. 2012, 21, 77–83. [Google Scholar]
- Little, D.G.; Song, K.M.; Katz, D.; Herring, J.A. Relationship of peak height velocity to other maturity indicators in idiopathic scoliosis in girls. J. Bone Jt. Surg. Am. 2000, 82, 685–693. [Google Scholar]
- Guo, X.; Chau, W.W.; Chan, Y.L.; Cheng, J.C. Relative anterior spinal overgrowth in adolescent idiopathic scoliosis. Results of disproportionate endochondral-membranous bone growth. J. Bone Jt. Surg. Br. 2003, 85, 1026–1031. [Google Scholar]
- Chu, W.C.; Lam, W.W.; Chan, Y.L.; Ng, B.K.; Lam, T.P.; Lee, K.M.; Guo, X.; Cheng, J.C. Relative shortening and functional tethering of spinal cord in adolescent idiopathic scoliosis?: Study with multiplanar reformat magnetic resonance imaging and somatosensory evoked potential. Spine 2006, 31, E19–E25. [Google Scholar]
- Zhu, F.; Qiu, Y.; Yeung, H.Y.; Lee, K.M.; Cheng, J.C.Y. Histomorphometric study of the spinal growth plates in idiopathic scoliosis and congenital scoliosis. Pediatr. Int. 2006, 48, 591–598. [Google Scholar]
- Dubousset, J.; Queneau, P.; Thillard, M.J. Experimental scoliosis induced by pieal and diencephalic leasions in young chickens: Its relation with clinical findings in idiopathic scoliosis. Orthop. Trans. 1983, 7, 7. [Google Scholar]
- Fjelldal, P.G.; Grotmol, S.; Kryvi, H.; Gjerdet, N.R.; Taranger, G.L.; Hansen, T.; Porter, M.J.; Totland, G.K. Pinealectomy induces malformation of the spine and reduces the mechanical strength of the vertebrae in atlantic salmon, salmo salar. J. Pineal Res. 2004, 36, 132–139. [Google Scholar]
- Machida, M.; Dubousset, J.; Yamada, T.; Kimura, J.; Saito, M.; Shiraishi, T.; Yamagishi, M. Experimental scoliosis in melatonin-deficient c57bl/6j mice without pinealectomy. J. Pineal Res. 2006, 41, 1–7. [Google Scholar]
- Bagnall, K.M.; Raso, V.J.; Hill, D.L.; Moreau, M.; Mahood, J.K.; Jiang, H.; Russell, G.; Bering, M.; Buzzell, G.R. Melatonin levels in idiopathic scoliosis. Diurnal and nocturnal serum melatonin levels in girls with adolescent idiopathic scoliosis. Spine 1996, 21, 1974–1978. [Google Scholar]
- Brodner, W.; Krepler, P.; Nicolakis, M.; Langer, M.; Kaider, A.; Lack, W.; Waldhauser, F. Melatonin and adolescent idiopathic scoliosis. J. Bone Jt. Surg. Br. 2000, 82, 399–403. [Google Scholar]
- Sadat-Ali, M.; Al-Habdan, I.; Al-Othman, A. Adolescent idiopathic scoliosis. Is low melatonin a cause? Jt. Bone Spine 2000, 67, 62–64. [Google Scholar]
- Moreau, A.; Wang, D.S.; Forget, S.; Azeddine, B.; Angeloni, D.; Fraschini, F.; Labelle, H.; Poitras, B.; Rivard, C.H.; Grimard, G. Melatonin signaling dysfunction in adolescent idiopathic scoliosis. Spine 2004, 29, 1772–1781. [Google Scholar]
- Moreau, A.; Akoume Ndong, M.Y.; Azeddine, B.; Franco, A.; Rompre, P.H.; Roy-Gagnon, M.H.; Turgeon, I.; Wang, D.; Bagnall, K.M.; Poitras, B.; et al. Molecular and genetic aspects of idiopathic scoliosis: Blood test for idiopathic scoliosis. Orthopade 2009, 38, 114–121. [Google Scholar]
- Azeddine, B.; Letellier, K.; Wang da, S.; Moldovan, F.; Moreau, A. Molecular determinants of melatonin signaling dysfunction in adolescent idiopathic scoliosis. Clin. Orthop. Relat. Res. 2007, 462, 45–52. [Google Scholar]
- Qiu, X.S.; Tang, N.L.; Yeung, H.Y.; Lee, K.M.; Hung, V.W.; Ng, B.K.; Ma, S.L.; Kwok, R.H.; Qin, L.; Qiu, Y.; et al. Melatonin receptor 1b (MTNR1B) gene polymorphism is associated with the occurrence of adolescent idiopathic scoliosis. Spine 2007, 32, 1748–1753. [Google Scholar]
- Takahashi, Y.; Matsumoto, M.; Karasugi, T.; Watanabe, K.; Chiba, K.; Kawakami, N.; Tsuji, T.; Uno, K.; Suzuki, T.; Ito, M.; et al. Lack of association between adolescent idiopathic scoliosis and previously reported single nucleotide polymorphisms in MATN1, MTNR1B, TPH1, and IGF1 in a Japanese population. J. Orthop. Res. 2011, 29, 1055–1058. [Google Scholar]
- Nelson, L.M.; Ward, K.; Ogilvie, J.W. Genetic variants in melatonin synthesis and signaling pathway are not associated with adolescent idiopathic scoliosis. Spine 2011, 36, 37–40. [Google Scholar]
- Morocz, M.; Czibula, A.; Grozer, Z.B.; Szecsenyi, A.; Almos, P.Z.; Rasko, I.; Illes, T. Association study of BMP4, IL6, LEPTIN, MMP3, and MTNR1B gene promoter polymorphisms and adolescent idiopathic scoliosis. Spine 2011, 36, E123–E130. [Google Scholar]
- Shyy, W.; Wang, K.; Gurnett, C.A.; Dobbs, M.B.; Miller, N.H.; Wise, C.; Sheffield, V.C.; Morcuende, J.A. Evaluation of GPR50, HMEL-1B, and ROR-α melatonin-related receptors and the etiology of adolescent idiopathic scoliosis. J. Pediatr. Orthop. 2010, 30, 539–543. [Google Scholar]
- Man, G.C.; Wang, W.W.; Yeung, B.H.; Lee, S.K.; Ng, B.K.; Hung, W.Y.; Wong, J.H.; Ng, T.B.; Qiu, Y.; Cheng, J.C. Abnormal proliferation and differentiation of osteoblasts from girls with adolescent idiopathic scoliosis to melatonin. J. Pineal Res. 2010, 49, 69–77. [Google Scholar]
- Man, G.C.; Wong, J.H.; Wang, W.W.; Sun, G.Q.; Yeung, B.H.; Ng, T.B.; Lee, S.K.; Ng, B.K.; Qiu, Y.; Cheng, J.C. Abnormal melatonin receptor 1b expression in osteoblasts from girls with adolescent idiopathic scoliosis. J. Pineal Res. 2011, 50, 395–402. [Google Scholar]
- Zhong, Z.M.; Li, T.; Xu, Z.X.; Meng, T.T.; Zeng, J.H.; Zheng, S.; Ye, W.B.; Wu, Q.; Chen, J.T. Effect of melatonin on the proliferation and differentiation of chondrocytes from rat vertebral body growth plate in vitro. Int. J. Med. Sci. 2013, 10, 1392–1398. [Google Scholar]
- Gao, W.; Lin, M.; Liang, A.; Zhang, L.; Chen, C.; Liang, G.; Xu, C.; Peng, Y.; Chen, C.; Huang, D.; et al. Melatonin enhances chondrogenic differentiation of human mesenchymal stem cells. J. Pineal Res. 2014, 56, 62–70. [Google Scholar]
- Aota, Y.; Terayama, H.; Saito, T.; Itoh, M. Pinealectomy in a broiler chicken model impairs endochondral ossification and induces rapid cancellous bone loss. Spine J. 2013, 13, 1607–1616. [Google Scholar]
- Bastepe, M.; Weinstein, L.S.; Ogata, N.; Kawaguchi, H.; Juppner, H.; Kronenberg, H.M.; Chung, U.I. Stimulatory g protein directly regulates hypertrophic differentiation of growth plate cartilage in vivo. Proc. Natl. Acad. Sci. USA 2004, 101, 14794–14799. [Google Scholar]
- Bi, W.; Deng, J.M.; Zhang, Z.; Behringer, R.R.; de Crombrugghe, B. Sox9 is required for cartilage formation. Nat. Genet. 1999, 22, 85–89. [Google Scholar]
- Huang, W.; Chung, U.I.; Kronenberg, H.M.; de Crombrugghe, B. The chondrogenic transcription factor Sox9 is a target of signaling by the parathyroid hormone-related peptide in the growth plate of endochondral bones. Proc. Natl. Acad. Sci. USA 2001, 98, 160–165. [Google Scholar]
- Peschke, E. Melatonin, endocrine pancreas and diabetes. J. Pineal Res. 2008, 44, 26–40. [Google Scholar]
- Sun, G.Q.; Wang, W.J.; Huang, A.B.; Cheng, J.C.; Qiu, Y. Expression of Melatonin Receptor in Chondrocyte of Adolescent Idiopathic Scoliosis; The International Research Society of Spinal Deformities: Liverpool, UK, 2008. [Google Scholar]
- Yim, A.P.; Yeung, H.Y.; Sun, G.; Lee, K.M.; Ng, T.B.; Lam, T.P.; Ng, B.K.; Qiu, Y.; Moreau, A.; Cheng, J.C. Abnormal skeletal growth in adolescent idiopathic scoliosis is associated with abnormal quantitative expression of melatonin receptor, mt2. Int. J. Mol. Sci. 2013, 14, 6345–6358. [Google Scholar]
- Zhang, L.; Zhang, J.; Ling, Y.; Chen, C.; Liang, A.; Peng, Y.; Chang, H.; Su, P.; Huang, D. Sustained release of melatonin from poly (lactic-co-glycolic acid) (PLGA) microspheres to induce osteogenesis of human mesenchymal stem cells in vitro. J. Pineal Res. 2013, 54, 24–32. [Google Scholar]
- Radio, N.M.; Doctor, J.S.; Witt-Enderby, P.A. Melatonin enhances alkaline phosphatase activity in differentiating human adult mesenchymal stem cells grown in osteogenic medium via MT2 melatonin receptors and the MEK/ERK (1/2) signaling cascade. J. Pineal Res. 2006, 40, 332–342. [Google Scholar]
- Satomura, K.; Tobiume, S.; Tokuyama, R.; Yamasaki, Y.; Kudoh, K.; Maeda, E.; Nagayama, M. Melatonin at pharmacological doses enhances human osteoblastic differentiation in vitro and promotes mouse cortical bone formation in vivo. J. Pineal Res. 2007, 42, 231–239. [Google Scholar]
- Nakade, O.; Koyama, H.; Ariji, H.; Yajima, A.; Kaku, T. Melatonin stimulates proliferation and type i collagen synthesis in human bone cells in vitro. J. Pineal Res. 1999, 27, 106–110. [Google Scholar]
- Roth, J.A.; Kim, B.G.; Lin, W.L.; Cho, M.I. Melatonin promotes osteoblast differentiation and bone formation. J. Biol. Chem. 1999, 274, 22041–22047. [Google Scholar]
- Liu, J.; Zhou, H.; Fan, W.; Dong, W.; Fu, S.; He, H.; Huang, F. Melatonin influences proliferation and differentiation of rat dental papilla cells in vitro and dentine formation in vivo by altering mitochondrial activity. J. Pineal Res. 2013, 54, 170–178. [Google Scholar]
- Burner, W.L.; Sherman, F.C. Osteoporosis and acquired back deformities. J. Pediatr. Orthop. 1982, 2, 383–385. [Google Scholar]
- Cheng, J.C.; Guo, X. Osteopenia in adolescent idiopathic scoliosis. A primary problem or secondary to the spinal deformity? Spine 1997, 22, 1716–1721. [Google Scholar]
- Cheng, J.C.; Guo, X.; Sher, A.H. Persistent osteopenia in adolescent idiopathic scoliosis. A longitudinal follow up study. Spine 1999, 24, 1218–1222. [Google Scholar]
- Yu, W.S.; Chan, K.Y.; Yu, F.W.; Yeung, H.Y.; Ng, B.K.; Lee, K.M.; Lam, T.P.; Cheng, J.C. Abnormal bone quality versus low bone mineral density in adolescent idiopathic scoliosis: A case-control study with in vivo high-resolution peripheral quantitative computed tomography. Spine J. 2013, 13, 1493–1499. [Google Scholar]
- Pandi-Perumal, S.R.; Srinivasan, V.; Maestroni, G.J.; Cardinali, D.P.; Poeggeler, B.; Hardeland, R. Melatonin: Nature’s most versatile biological signal? FEBS J. 2006, 273, 2813–2838. [Google Scholar]
- Lee, K.M.; Fung, K.P.; Leung, P.C.; Leung, K.S. Identification and characterization of various differentiative growth plate chondrocytes from porcine by countercurrent centrifugal elutriation. J. Cell Biochem. 1996, 60, 508–520. [Google Scholar]
- Hidvegi, N.C.; Sales, K.M.; Izadi, D.; Ong, J.; Kellam, P.; Eastwood, D.; Butler, P.E. A low temperature method of isolating normal human articular chondrocytes. Osteoarthr. Cartil. 2006, 14, 89–93. [Google Scholar]
- Dubocovich, M.L.; Masana, M.I.; Iacob, S.; Sauri, D.M. Melatonin receptor antagonists that differentiate between the human mel1a and mel1b recombinant subtypes are used to assess the pharmacological profile of the rabbit retina ml1 presynaptic heteroreceptor. Naunyn. Schmiedebergs Arch. Pharmacol. 1997, 355, 365–375. [Google Scholar]
- Rodriguez, L.; Cheng, Z.; Chen, T.H.; Tu, C.; Chang, W. Extracellular calcium and parathyroid hormone-related peptide signaling modulate the pace of growth plate chondrocyte differentiation. Endocrinology 2005, 146, 4597–4608. [Google Scholar]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Wang, W.W.-J.; Man, G.C.-W.; Wong, J.H.; Ng, T.-B.; Lee, K.-M.; Ng, B.K.-W.; Yeung, H.-Y.; Qiu, Y.; Cheng, J.C.-Y. Abnormal Response of the Proliferation and Differentiation of Growth Plate Chondrocytes to Melatonin in Adolescent Idiopathic Scoliosis. Int. J. Mol. Sci. 2014, 15, 17100-17114. https://doi.org/10.3390/ijms150917100
Wang WW-J, Man GC-W, Wong JH, Ng T-B, Lee K-M, Ng BK-W, Yeung H-Y, Qiu Y, Cheng JC-Y. Abnormal Response of the Proliferation and Differentiation of Growth Plate Chondrocytes to Melatonin in Adolescent Idiopathic Scoliosis. International Journal of Molecular Sciences. 2014; 15(9):17100-17114. https://doi.org/10.3390/ijms150917100
Chicago/Turabian StyleWang, William Wei-Jun, Gene Chi-Wai Man, Jack Ho Wong, Tzi-Bun Ng, Kwong-Man Lee, Bobby Kin-Wah Ng, Hiu-Yan Yeung, Yong Qiu, and Jack Chun-Yiu Cheng. 2014. "Abnormal Response of the Proliferation and Differentiation of Growth Plate Chondrocytes to Melatonin in Adolescent Idiopathic Scoliosis" International Journal of Molecular Sciences 15, no. 9: 17100-17114. https://doi.org/10.3390/ijms150917100