
Abstract
The etiologic factors in the development of autoimmune thyroid diseases (AITDs) are not fully understood. We investigated the role of skewed X-chromosome inactivation (XCI) mosaicism in female predisposition to AITDs. One hundred and ten female AITDs patients (81 Hashimoto's thyroiditis (HT), 29 Graves' disease (GD)), and 160 female controls were analyzed for the androgen receptor locus by the HpaII/polymerase chain reaction assay to assess XCI patterns in DNA extracted from peripheral blood cells. In addition, thyroid biopsy, buccal mucosa, and hair follicle specimens were obtained from five patients whose blood revealed an extremely skewed pattern of XCI, and the analysis was repeated. Skewed XCI was observed in DNA from peripheral blood cells in 28 of 83 informative patients (34%) as compared with 10 of 124 informative controls (8%, P<0.0001). Extreme skewing was present in 16 patients (19%), but only in three controls (2.4%, P<0.0001). The buccal mucosa, and although less marked, the thyroid specimens also showed skewing. Analysis of two familial cases showed that only the affected individuals demonstrate skewed XCI patterns. Based on these results, skewed XCI mosaicism may play a significant role in the pathogenesis of AITDs.
Similar content being viewed by others

Introduction
Hashimoto's thyroiditis (HD) and Graves' disease (GD) are autoimmune thyroid diseases associated with multiple genetic factors. Although the pathogenesis is poorly understood, a widely accepted model suggests an inherited background, which predisposes the subjects to autoimmunity. Additional intrinsic and extrinsic factors such as hormones and the environment may ultimately trigger or contribute to the development of the disease phenotype.1 Extensive linkage genome screens during the past decade have resulted in the identification of several thyroid-specific susceptibility genes and/or loci, but confirmation through multiple population studies is still awaited for the majority of these loci.1, 2 A common feature of autoimmune diseases, including autoimmune thyroid diseases (AITDs), is an increased prevalence in women when compared with men. The most striking sex differences are observed in AITDs, scleroderma, Sjögren's syndrome, and systemic lupus erythematosus, which are diseases where over 80% of the patients are females.3
It has been demonstrated that risk of autoimmunity could be increased by a lack of exposure to self-antigens in the thymus and the presence of autoreactive T cells.4, 5, 6 Disturbances in the X-chromosome inactivation (XCI) process provide a potential mechanism whereby the lack of exposure to self-antigens could occur,7, 8 including AITDs.9, 10 X-chromosome inactivation is a physiologic process that takes place in early female development and results in the transcriptional silencing of one of the pair of X chromosomes.11 As a result of this epigenetic regulation, a random inactivation of the X chromosome inherited from either parent occurs and normal female subjects are thus a mosaic of two cell populations. It is therefore an attractive hypothesis that skewed XCI could lead to the escape of X-linked self-antigens from presentation in the thymus or in other peripheral sites that are involved in tolerance induction, inadequate thymic deletion, and finally loss of T-cell tolerance. Indeed, we recently observed skewed XCI in blood cells of women with scleroderma.12
Based on our observation that an association exists between skewed XCI and female predisposition to autoimmunity, we hypothesized that skewed XCI may be involved in the pathogenesis of AITDs, particularly in the hematopoietic compartment. We observed extremely skewed XCI in the blood samples of a significant proportion of female patients with AITDs.
Methods
Patients and pedigree analysis
Caucasian women diagnosed with AITDs (n=110), and healthy female controls with no history of autoimmune disease and cancer (n=160) were included in the study. Among the patients, 81 were diagnosed with HT and 29 with GD. The mean ages were 44.8±14.1 (mean±SD) years for AITDs (46±14.2 years in the Hashimoto patients, and 40.6±13.2 years in the Graves' patients), and 46±10 for controls. The duration of the symptoms was 5.7±7.4 years among the AITDs patients (5.7±7 years in the Hashimoto patients and 6±8.5 years in the Graves' patients). The mean age of diagnosis was 39±12 years. All of the patients had attended the outpatient clinics of the Endocrinology and Metabolic Diseases Department of Ankara University School of Medicine for at least 1 year since the onset of disease. Patients were randomly chosen for the study.
All clinical investigations described in this manuscript were conducted in accordance with the guidelines in the Declaration of Helsinki (http://www.wma.net). The ethics review board of the participating institutions approved the study protocol. Informed consent was obtained from all subjects.
The diagnosis of HD was made by the existence of a firm goitre in combination with elevated thyroid auto-antibodies (thyroglobulin and/or thyroid peroxidase), a low ultrasonographic echogenity of the gland, and demonstration of lymphocytic infiltration by fine-needle aspiration biopsy and/or biochemical hypothyroidism. The diagnosis of GD was based on biochemical hyperthyroidism, and a diffuse symmetrical goitre in combination with positive thyroid antibodies (thyroglobulin, thyroid peroxidase or TSH receptor). In addition, thyroid ophthalmopathy and/or diffuse hyperplasia on an isotope scan or ultrasonography demonstrating homogenous echo texture may accompany the clinical picture.
Following the XCI studies, a complete pedigree analysis was carried out for 64 individuals informative for the AR polymorphism with medical follow-up of reported AITDs among family members when possible. Owing to emigration or unwillingness to contribute family information, data could not be obtained from the remaining 19 participants. Family history of AITDs was determined by reviewing the probands' pedigree to determine the number of relatives affected by these autoimmune diseases. Only first- and second-degree relatives were counted. A positive family history was noted if one additional AITD was documented by medical review.
X-chromosome inactivation analysis
Genotyping of a highly polymorphic CAG repeat in the androgen-receptor (AR) gene was performed to assess the XCI patterns as described elsewhere.12, 13 Densitometric analysis of the alleles was performed at least twice for each sample using the MultiAnalyst version 1.1 software. A corrected ratio (CrR) was calculated by dividing the ratio of the predigested sample (upper/lower allele) by the ratio of the nonpredigested sample for normalization of the ratios that were obtained from the densitometric analyses. The use of CrR compensates for preferential amplification of the shorter allele when the number of PCR cycles increases.14 A skewed population is defined as a cell population with greater than 80% expression of one of the AR alleles. This corresponds to CrR values of <0.33 or >3.
Haplotype analysis
Human MapPairs Version 10 purchased from Research Genetics (currently available by Invitrogen, CA, USA) was used to screen the X chromosome. Site-specific PCR, 6% polyacrylamide gel electrophoresis, and silver staining techniques were used for genotyping the individuals. Gels were manually pictured and genotyped. Cyrillic program (version 2) was used to generate the haplotypes. A total of 27 X-chromosome-specific DNA markers from the MapPairs Panel were genotyped. Map order and physical positions (Mb) of the additional polymorphic DNA markers were obtained from USCS genome browser (The University of California Santa Cruz, CA, USA http://genome.ucsc.edu/).
Statistical methods
The results from control and test groups in XCI studies were compared by χ2 test with Yate's correction.
Results
PCR-based X-inactivation study of peripheral blood
XCI status was informative in 83 of the 110 AITDs patients and in 124 of the 160 controls. Some heterozygous individuals were considered uninformative since only those whose alleles resolve adequately for densitometric analyses were included in the study. Skewed XCI (>80% skewing) was observed in 28 of the 83 patients (34%), and 10 of the 124 controls (8%) (P<0.0001). When the data for the two groups of AITDs patients was analyzed independently, 23/67 (34.33%, P<0.0001) of the Hashimoto's patients and 5/16 (31.25%, P=0.0167) of the Graves' patients were found to display the skewed XCI in blood. More importantly, extremely skewed XCI, defined as >90% inactivation of one allele, was present in 16 patients (19%), and in only three controls (2.4%, P<0.0001) (see Table 1). Extremely skewed XCI is a rare event in the general population. It has been reported in only 1–2% of women aged 20–40 years, and in 2–4% of women aged 55–72 years.15, 16 The distribution of XCI skewing in the general population is thought to be mainly due to chance deviations from 50:50 as a result of the limited number of embryonic cells present (4–20) at the time of XCI.17 Age alone is unlikely to influence the strikingly bimodal data in our AITDs patients (Figure 1). We did not observe a shift towards the skewed range in older patients and controls.

Distribution of X-inactivation patterns according to age in AITDs patients and control subjects.
PCR-based X-inactivation study of thyroid biopsy, buccal mucosa, and hair follicle specimens
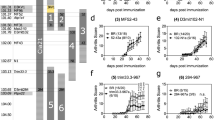
Thyroid biopsy, buccal mucosa, and hair follicle specimens were obtained from five patients (04-121, 04-198, 04-214, 04-221, and 04-225). Their blood XCI profile displayed almost exclusive representation of only one allele of the AR polymorphism in their methylation-sensitive PCR assay, which indicates extremely skewed XCI. Five randomly selected patients showed skewing in the same direction for all tissues, except hair follicle, that in the thyroid being less marked than blood and buccal cells (Figure 2). Hair follicle specimens had a random XCI pattern. The allele ratios are given in Table 2.

X-inactivation analysis of androgen receptor locus. PCR products of undigested (−) and HpaII-digested (+) DNA from peripheral blood, thyroid biopsy, buccal, and hair follicle samples of AITDs patients 04-121, 04-198, 04-214, 04-221, and 04-225 are shown. Two alleles are seen in undigested samples, whereas a single allele resulting from extremely skewed XCI is clearly visible in all peripheral blood samples. Allele ratios are given in the text and in Table 2. M: marker (pUC mix 8), 331 and 242 bp fragments are visible.
Pregnancy history and pedigree analysis
Characteristics of the AITDs patients with skewed and random XCI are shown in Table 3. Only those patients with a complete pregnancy and family history are included in this table. The pedigrees of many AITDs probands with skewed XCI versus those with random XCI were interesting in two aspects. First, recurrent spontaneous abortions (defined as three or more pregnancy losses), which have been shown to be associated with skewed XCI,16, 18 occurred in four of 25 (16%) of our AITDs probands with skewed XCI. Conversely, a history of recurrent spontaneous abortions was negative both in the patients with random XCI and in the control group subjects (P<0.0199). Although the etiology of recurrent abortions in thyroid autoimmunity remains unknown, women who present with thyroid antibodies in the first trimester of pregnancy have a two- to four-fold increase in their miscarriage rates.19
Second, a positive family history, particularly in the skewed group, was apparent (12/25, 48% in the skewed; and 10/39, 25.6% in the random groups). We therefore contacted all of the 12 probands in an attempt to extend the X-chromosome inactivation studies to other family members. Initially, a positive response was received from three families, but blood samples could be obtained from the family members of only two probands (04-445, Family 1; and 04-298, Family 2). An important observation emerges from a study of these families: only the affected individuals demonstrate skewed XCI patterns. For example, XCI is extremely skewed in the affected sister and mother of 04-445 (Family 1), but random in the two unaffected sisters. The inactive X chromosome here is of maternal origin. In patient 04-298 (Family 2), skewing in the 80–89% range is noted for her affected sister, but unfortunately her mother was not informative for the AR polymorphism. Interestingly, the inactive X chromosome appears to be of paternal origin in Family 2 (Supplementary Figures 1 and 2).
Haplotype analysis
Because XCI segregates as a heritable trait associated with the disease in two generations of Family 1, we performed haplotype analysis by using polymorphic X-chromosomal markers to determine possible segregation between the disease and marker alleles. Although the size of this family is not large enough to prove linkage, it still provides valuable information about the exclusion area on the X chromosome. This helps to define a minimal critical region on the X chromosome, which might be associated with AITDs. Xp11-q13 (GATA144DO4, DXS7132, and AR) and Xp22 DNA markers (DXS8022, DXS987, and DX9902) showed concordance among the affected individuals indicating positive segregation between the disease and marker alleles. The haplotype structure is shown in Figure 3. However, lod score20 analysis did not allow formal acceptance of linkage to any loci mainly due to the small size of the family.

Haplotype structure of Family 1. Patient 04-445 was arbitrarily selected to construct the haplotype. Maternally inherited haplotype was highlighted with solid black bar. Haplotypes of the remaining sibs were compared with the reference individual (04-445), and shared portions were also marked with solid bars. Noninformativeness in the crossover regions were demonstrated with thin bars. The regions between the DNA markers DXS8051 and DXS8036 as well as DXS8014 and AR regions on Xp22 and Xp11-q13 regions, respectively, were not excluded since positive segregation between the disease and marker alleles was observed.
Discussion
The autoimmune diseases include more than 70 chronic disorders that affect approximately 5% of the population. A reduction in sex ratio (male:female) is characteristic of most such diseases, including AITDs.3 Even though the female prevalence of autoimmune diseases has been recognized for over a hundred years, candidate mechanisms that could be important in pathogenesis have been uncovered only during the past two decades. These include genetic traits associated with autoimmunity,21 pregnancy-related microchimerism,22 and disturbances in XCI mosaicism in female subjects.12 In this study, we demonstrate skewed XCI patterns in peripheral blood mononuclear cells of a significant proportion (34%) of female subjects with AITDs. Approximately 8% of female control subjects demonstrate skewed X-inactivation patterns ⩾80:20, which is consistent with previous estimates.16, 18, 23 The effect is more pronounced at patterns of X-inactivation ⩾90:10; nearly 20% of AITDs patients show such skewing (Supplementary Figure 3), compared with only a few percent of female control subjects. Our results show that factors associated with extremely skewed XCI could account for a significant proportion of female patients with AITDs.
Skewed XCI is a result of primary or secondary causes. The former is bias in the initial choice of which X chromosome is inactivated due to germline XIST (X-inactive-specific transcript) mutations.24 The secondary causes are deleterious X-linked mutations, X chromosome rearrangements, aging, twinning, or monoclonal expansion of cells (for a review, see Brown25). We believe that deleterious X-linked mutations or X chromosome rearrangements and their differential expression patterns could provide a disadvantage to blood and buccal cells, and possibly to thyroid cells in AITDs patients, and lead to skewed XCI. This has been supported by our observation that maternally inherited skewed XCI profile accompanies the disease phenotype for our AITDs Family 1. We observed segregation between the disease and marker alleles with the DNA markers residing on the distal short arm and pericentromeric regions of the X chromosome in this family. Although examples of skewed X-inactivation segregating with a trait have been reported previously,18, 26 this is the first example in AITDs to the best of our knowledge. In a recently published study on a three-generation kindred, extreme skewing of X inactivation was documented in three female subjects who have hemophilia A.26 Since the inactive X was always of paternal origin in affected female subjects, the authors concluded that skewing in the family resulted from an abnormality in the initial choice process. This prevented the X chromosome, which carried the mutant FVIII allele, from being an inactive X. In our Family 2 with two affected sisters, the inactive X chromosome was of paternal origin like the hemophilia A family. Extension of both the XCI and linkage studies to large cohorts with familial AITDs cases could prove to be very rewarding in understanding the relation between skewed XCI and autoimmune thyroidites.
Studies that aim to delineate the medical consequences of skewed X-inactivation have shown that clinical manifestation of X-linked disorders in female subjects could be influenced by disturbances in the XCI process.27 In addition, it has been hypothesized that skewed XCI could be a factor that influences female predisposition to autoimmunity.7, 8 Now that we have demonstrated skewed patterns of XCI in a significant proportion of female AITD patients, deviation from the physiological range of XCI mosaicism could be considered as a potential mechanism contributing to disease pathogenesis. This is further supported by the recently reported observation that female twins with AITDs have a high frequency of skewed XCI.28
Although extremely skewed XCI is rare, it does not always lead to the development of AITDs. A subsequent event, such as environmental exposure to viral, chemical, or other agents may trigger a cascade that results in AITDs. In addition, the co-inheritance of genetic susceptibility factors, such as functional variants in vital negative regulatory molecules of the immune system,29, 30 may exacerbate the effects of skewed XCI and contribute to the development of autoimmune diseases including AITDs.
References
Ban Y, Tomer Y : Susceptibility genes in thyroid autoimmunity. Clin Dev Immunol 2005; 12: 47–58.
Dechairo BM, Zabaneh D, Collins J et al: Association of the TSHR gene with Graves' disease: the first disease specific locus. Eur J Hum Genet 2005; 13: 1223–1230.
Whitacre CC : Sex difference in autoimmune disease. Nat Immunol 2001; 2: 777–780.
Laufer TM, DeKoning J, Markowitz JS, Lo D, Glimcher LH : Unopposed positive selection and autoreactivity in mice expressing class II MHC only on thymic cortex. Nature 1996; 383: 81–85.
Klein L, Klugmann M, Nave KA, Tuohy VK, Kyewski B : Shaping of the autoreactive T-cell repertoire by a splice variant of self protein expressed in thymic epithelial cells. Nat Med 2000; 6: 56–61.
Kyewski B, Derbinski J : Self-representation in the thymus: an extended view. Nat Rev Immunol 2004; 4: 688–698.
Kast RE : Predominance of autoimmune and rheumatic diseases in females. J Rheumatol 1977; 4: 288–292.
Stewart JJ : The female X-inactivation mosaic in systemic lupus erythematosus. Immunol Today 1998; 19: 352–357.
Barbesino G, Tomer Y, Concepcion ES, Davies TF, Greenberg DA : Linkage analysis of candidate genes in autoimmune thyroid disease. II. Selected gender-related genes and the X-chromosome. International Consortium for the Genetics of Autoimmune Thyroid Disease. J Clin Endocrinol Metab 1998; 83: 3290–3295.
Tomer Y, Barbesino G, Greenberg DA, Concepcion E, Davies TF : Mapping the major susceptibility loci for familial Graves' and Hashimoto's diseases: evidence for genetic heterogeneity and gene interactions. J Clin Endocrinol Metab 1999; 84: 4656–4664.
Lyon MF : Gene action in the X-chromosome of the mouse (Mus musculus L). Nature 1961; 190: 372–373.
Ozbalkan Z, Bagislar S, Kiraz S et al: Skewed X chromosome inactivation in blood cells of women with scleroderma. Arthritis Rheum 2005; 52: 1564–1570.
Allen RC, Zoghbi HY, Moseley AB, Rosenblatt HM, Belmont JW : Methylation of HpaII and HhaI sites near the polymorphic CAG repeat in the human androgen-receptor gene correlates with X chromosome inactivation. Am J Hum Genet 1992; 51: 1229–1239.
Delforge M, Demuynck H, Vandenberghe P et al: Polyclonal primitive hematopoietic progenitors can be detected in mobilized peripheral blood from patients with high-risk myelodysplastic syndromes. Blood 1995; 86: 3660–3667.
Buller RE, Sood AK, Lallas T, Buekers T, Skilling JS : Association between nonrandom X-chromosome inactivation and BRCA1 mutation in germline DNA of patients with ovarian cancer. J Natl Cancer Inst 1999; 91: 339–346.
Sangha KK, Stephenson MD, Brown CJ, Robinson WP : Extremely skewed X-chromosome inactivation is increased in women with recurrent spontaneous abortion. Am J Hum Genet 1999; 65: 913–917.
Monteiro JDC, Vlietinck R, Kohn N, Lesser M, Gregersen PK : Commitment to X inactivation procedes the twining event in monochorionic MZ twins. Am J Hum Genet 1998; 63: 339–346.
Pegoraro E, Whitaker J, Mowery-Rushton P, Surti U, Lanasa M, Hoffman EP : Familial skewed X inactivation: a molecular trait associated with high spontaneous-abortion rate maps to Xq28. Am J Hum Genet 1997; 61: 160–170.
Stagnaro-Green A : Thyroid autoimmunity and the risk of miscarriage. Best Pract Res Clin Endocrinol Metab 2004; 18: 167–181.
Abecasis GR, Cherny SS, Cookson WO, Cardon LR : Merlin-rapid analysis of dense genetic maps using sparse gene flow trees. Nat Genet 2002; 30: 97–101.
Rioux JD, Abbas AK : Paths to understanding the genetic basis of autoimmune disease. Nature 2005; 435: 584–589.
Adams KM, Nelson JL : Microchimerism: an investigative frontier in autoimmunity and transplantation. JAMA 2004; 291: 1127–1131.
Chitnis S, Monteiro J, Glass D et al: The role of X-chromosome inactivation in female predisposition to autoimmunity. Arthritis Res 2000; 2: 399–406.
Puck J, Willard H : X-inactivation in females with X-linked disease. N Engl J Med 1998; 338: 325–328.
Brown CJ : Skewed X-chromosome inactivation: cause or consequence? J Natl Cancer Inst 1999; 91: 304–305.
Bicocchi MP, Migeon BR, Pasino M et al: Familial nonrandom inactivation linked to the X inactivation centre in heterozygotes manifesting haemophilia A. Eur J Hum Genet 2005; 13: 635–640.
Lyon MF : X-chromosome inactivation and human genetic disease. Acta Paediatr Suppl 2002; 91: 107–112.
Brix TH, Knudsen GP, Kristiansen M, Kyvik KO, Ørstavik KH, Hegedüs L : High frequency of skewed X chromosome inactivation in females with autoimmune thyroid disease. A possible explanation for the female predisposition to thyroid autoimmunity. J Clin Endocrinol Metab 2005; 90: 5949–5953.
Ueda H, Howson JM, Esposito L et al: Association of the T-cell regulatory gene CTLA4 with susceptibility to autoimmune disease. Nature 2003; 423: 506–511.
Criswell LA, Pfeiffer KA, Lum RF et al: Analysis of families in the multiple autoimmune disease genetics consortium (MADGC) collection: the PTPN22 620W allele associates with multiple autoimmune phenotypes. Am J Hum Genet 2005; 76: 261–271.
Acknowledgements
We thank Margaret Sands, Iclal Ozcelik, and Ozlen Konu for critical reading of the manuscript. This study was supported by grants from the Scientific and Technical Research Council of Turkey – TUBITAK-SBAG 2513, International Centre for Genetic Engineering and Biotechnology – ICGEB-CRP/TUR04-01, and Bilkent University Research Fund (to Dr Ozcelik).
Author information
Authors and Affiliations
Corresponding author
Additional information
Supplementary Information accompanies the paper on European Journal of Human Genetics website (http://www.nature.com/ejhg)
Rights and permissions
About this article
Cite this article
Ozcelik, T., Uz, E., Akyerli, C. et al. Evidence from autoimmune thyroiditis of skewed X-chromosome inactivation in female predisposition to autoimmunity. Eur J Hum Genet 14, 791–797 (2006). https://doi.org/10.1038/sj.ejhg.5201614
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/sj.ejhg.5201614
Keywords
This article is cited by
-
X-linked genes exhibit miR6891-5p-regulated skewing in Sjögren’s syndrome
Journal of Molecular Medicine (2022)
-
Sex-related factors in autoimmune liver diseases
Seminars in Immunopathology (2019)
-
Sex differences in the expression of lupus-associated miRNAs in splenocytes from lupus-prone NZB/WF1 mice
Biology of Sex Differences (2013)
-
Skewed X-chromosome inactivation in patients with esophageal carcinoma
Diagnostic Pathology (2013)
-
Mechanisms and Pathophysiology of Autoimmune Disease
Clinical Reviews in Allergy & Immunology (2012)

{kind=link}
{kind=link}
{kind=link}