Article Text

Abstract
Background Hereditary myopathy with lactic acidosis and myopathy with deficiency of succinate dehydrogenase and aconitase are variants of a recessive disorder characterised by childhood-onset early fatigue, dyspnoea and palpitations on trivial exercise. The disease is non-progressive, but life-threatening episodes of widespread weakness, metabolic acidosis and rhabdomyolysis may occur. So far, this disease has been molecularly defined only in Swedish patients, all homozygous for a deep intronic splicing affecting mutation in ISCU encoding a scaffold protein for the assembly of iron–sulfur (Fe-S) clusters. A single Scandinavian family was identified with a different mutation, a missense change in compound heterozygosity with the common intronic mutation. The aim of the study was to identify the genetic defect in our proband.
Methods A next-generation sequencing (NGS) approach was carried out on an Italian male who presented in childhood with ptosis, severe muscle weakness and exercise intolerance. His disease was slowly progressive, with partial recovery between episodes. Patient’s specimens and yeast models were investigated.
Results Histochemical and biochemical analyses on muscle biopsy showed multiple defects affecting mitochondrial respiratory chain complexes. We identified a single heterozygous mutation p.Gly96Val in ISCU, which was absent in DNA from his parents indicating a possible de novo dominant effect in the patient. Patient fibroblasts showed normal levels of ISCU protein and a few variably affected Fe-S cluster-dependent enzymes. Yeast studies confirmed both pathogenicity and dominance of the identified missense mutation.
Conclusion We describe the first heterozygous dominant mutation in ISCU which results in a phenotype reminiscent of the recessive disease previously reported.
- Iscu
- Mitochondrial Myopathy
- De Novo Mutation
- Fe-s Cluster
This is an Open Access article distributed in accordance with the terms of the Creative Commons Attribution (CC BY 4.0) license, which permits others to distribute, remix, adapt and build upon this work, for commercial use, provided the original work is properly cited. See: http://creativecommons.org/licenses/by/4.0/
Statistics from Altmetric.com
Introduction
Iron–sulfur (Fe-S) clusters are prosthetic groups found in several mitochondrial, cytosolic and nuclear enzymes, which play a role in fundamental cellular processes, such as respiration, DNA synthesis and repair, ribosome biogenesis and iron metabolism. In eukaryotes, the biogenesis of Fe-S clusters is performed by two main multiprotein machineries, the ISC (iron–sulfur cluster assembly) machinery localised in mitochondria and the CIA (cytosolic iron–sulfur protein assembly) machinery localised in the cytosol.1 2 Fe-S clusters are found in almost all living organisms, and the most common stoichiometric species include [2Fe-2S], [3Fe-4S] and [4Fe-4S] structures in which the Fe ions are co-ordinated, for example, by cysteine thiol groups or histidine residues.
The ISC machinery is involved in the biogenesis of Fe-S proteins in mitochondria enzymes as well as in the cytosol and nucleus. The current understanding of the molecular mechanism of mitochondrial Fe-S protein biogenesis has been worked out in both yeast and human cells. The ISC machinery comprises 18 known proteins that perform several steps of Fe-S cluster synthesis, transfer and insertion into recipient proteins including subunits of the mitochondrial respiratory chain (MRC) complexes I, II and III, mitochondrial aconitase (mACO) and lipoic acid synthase (LIAS).3 De novo synthesis of the [2Fe-2S] cluster is accomplished on the scaffold protein ISCU. This reaction requires the cysteine desulfurase NFS1 with its stabilising partners ISD11/LYRM4 and ACP1, frataxin/FXN as an iron donor and/or regulator of cysteine desulfurase activity, and the ferredoxin FDX2 (MIM 614585) as an electron donor for sulfur reduction.4 5 Conflicting results have been published regarding the role of FDX1 (MIM 103260) in this process.6 7 All these ISC proteins form a dynamic complex with ISCU. Dissociation of the preformed Fe-S cluster from the ISCU scaffold and transfer to intermediate carriers, for example the monothiol glutaredoxin GLRX5 (MIM 609588), is mediated by a dedicated Hsp70-Hsp40 chaperone system (HSC20/HSP70).8 The transiently GLRX5-bound [2Fe-2S] cluster is inserted into [2Fe-2S] targets or used for [4Fe-4S] cluster synthesis by ISCA1-ISCA2-IBA57. Finally, the cluster is inserted into target apoproteins with the help of factors including IND1 (MIM 613621), NFU1 (MIM 608100) and BOLA3 (MIM 613183). A different targeting mode has been suggested, based on interactions of the adaptable HSC20/HSPA9 scaffold complex with LYR motifs of SDHAF1 for complex II,9 with LYRM7 for complex III or directly with Fe-S cluster subunits of complex I.10
Recessive mutations in ISCU have been described in patients presenting myopathy with severe exercise intolerance and myoglobinuria (MIM 255125). A homozygous intronic transversion (c.418+382G>C or IVS5 +382G>C) was initially reported in patients from northern Sweden, with associated deficiencies of succinate dehydrogenase and aconitase in skeletal muscle.11 12 The mutation causes the retention of an ‘extra exon’, leading to marked reduction of ISCU mRNA and protein in patient muscle. The splicing defect was shown to be selective for muscle tissue, thus explaining the muscle-specific phenotype of this disorder.13 Later, compound heterozygosity for the common intronic mutation and a missense c.149G>A/p.G50E substitution was found in two brothers with Swedish/Finnish origin. These boys had a more severe phenotype than patients homozygous for the intronic mutation, with progressive and severe muscle weakness, muscle wasting and heart involvement.14
Contrary to the muscular phenotype of ISCU mutant patients, mutations in other components of the core Fe-S assembly complex cause neurological diseases (eg, Friedreich’s ataxia, MIM 229300, due to FXN mutations) or multisystem disorders (eg, combined oxidative phosphorylation deficiency 19, MIM 615595, due to ISD11/LYRM4 mutations).
We report here a patient with myopathy, lactic acidosis and combined MRC complex deficiency, caused by a de novo heterozygous missense pathological variant in ISCU.
Methods
Histochemical and biochemical studies in skeletal muscle
Muscle morphology and histochemistry, respiratory chain activities of complexes I to IV and pyruvate dehydrogenase complex (PDHC) assays were performed as previously described.15–17 Histochemical staining of iron using Prussian blue colour was performed as previously described.18
Genetic analysis
Genomic DNA was extracted from peripheral blood by standard methods. Whole exome sequencing (WES) and variants filtering were performed as previously described.19 Variants identified by WES were validated by Sanger sequencing. For deep sequencing of parental DNAs, the PCR products were processed with Nextera XT DNA sample preparation kit (Illumina). Sequencing was performed on an Illumina MiSeq instrument.
RNA was extracted from skin fibroblasts, and 1 µg was used as template for RT-PCR to obtain full-length cDNA. ISCU transcript was amplified by PCR and run through a 1% agarose gel in order to detect potential splicing alterations. PCR products were also sequenced in order to confirm genomic variants and unmask potential events of nonsense-mediated decay.
Cell culture and biochemical analysis of fibroblast samples
Fibroblasts obtained from skin biopsy were grown in 1 g/L glucose DMEM-F14 (Euroclone) supplemented with 20% fetal bovine serum (FBS), 1% uridine, 1% L-glutamine and 0.2% sodium pyruvate.
For enzyme activity measurements, cells were treated with digitonin in order to separate a cytosolic cell fraction from a crude mitochondria-containing organellar fraction.20 Biochemical assays were essentially performed as described.16 Analysis of steady-state protein levels by immunoblotting was carried out by common methods, using a 6%–20% sodium dodecyl sulfate polyacrylamide (SDS-PA) gradient gel. Blocked membranes were probed with primary antibodies (online supplementary table S1), and antigens were visualised by horseradish peroxidase (HRP) coupled secondary reagents and a chemiluminescence reaction.
Yeast studies
Details on yeast strains, media, cloning procedures and vectors21 22 as well as on generation of mutant allele and construction of mutant strains23–25 are reported in the online supplementary data
Complex II (succinate dehydrogenase (SDH)) and complex IV (cytochrome c oxidase (COX)) specific activities were measured on a mitochondrial-enriched fraction prepared as previously described.26 27 Aconitase activity was measured in whole-cell extracts.28 In vivo radiolabelling of yeast cells with 55FeCl3 (ICN) and measurement of 55Fe-incorporation into Fe-S proteins by immunoprecipitation and scintillation counting were performed as described.29 Antibodies against c-Myc were obtained from Santa-Cruz. The green fluorescent protein (GFP) based reporter assay for determination of FET3 promoter strength was described previously.29 The iron content was determined by a colorimetric assay, essentially as described before.30 31
Bioinformatics and structural analysis tools
The effect of the p.(Gly96Val) substitution on ISCU function was predicted using the in silico tools (ie, SIFT, Polyphen2 and EVmutation), all running recommended parameters. All images of the Escherichia coli IscU-IscS (PDB ID:3LVL) complex were made by using Visual Molecular Dynamics viewer.32 The evaluation of the impact of the mutated residue and the protein–protein interaction analysis were performed by using Swiss PDB viewer,33 and FirstGlance in Jmol (http://www.jmol.org)
Results
Case report
The patient is a 23-year-old Italian male, first child from non-consanguineous parents, born at term after a normal pregnancy by caesarean delivery. He has a healthy younger brother. He started walking at 18 months, but he always presented some walking difficulties, with frequent falls. Parents reported easy fatigability since the first years of life. At 7 years of age, the neurological examination showed bilateral ptosis not associated with ophthalmoparesis, muscle hypotonia and wasting, and absent deep tendon reflexes. No obvious weakness was present, but marked exercise intolerance was reported. Neither cognitive impairment nor other central nervous system (CNS) involvements were noticed. The brain nuclear magnetic resonance (NMR) and the EEG were normal. Electromyography showed myopathic changes in all tested muscles. Creatine kinase (CK) level was slightly increased (about 300 U/L; normal values, n.v.:<180), lactate acid was high in blood (4.6 mmol/L; n.v.: 0.4–2.2). He showed leucopenia (2.94×103/µL; n.v.: 5–14) and anaemia (haemoglobin 10.9 g/dL; n.v. 13–16) with microcythemia (72.9; n.v. 80–99); a bone marrow biopsy performed at 7 years was normal. No heart involvement was present: echocardiogram and ECG were both normal. Over time, his clinical conditions worsened and he started presenting episodes of profound exercise intolerance and weakness, with partial recovery of muscle weakness between episodes in about 2 weeks. During these episodes, the patient was unable to walk, and showed tachycardia; neither breathing shortness nor dysphagia was noticed. At the neurological examination at 17 years of age, he presented predominantly distal limb weakness with muscle hypotrophy; deep tendon reflexes were absent. He was able to stand up from a chair and walk, but was unable to run. No signs of CNS involvement were present; CK level remained high (about 1700 U/L). The disease has since been slowly progressive, punctuated by episodes of acute weakness, with preserved cognitive function and no other signs of CNS involvement.
Histochemical and biochemical analyses in skeletal muscle
A first muscle biopsy was performed at 8 years of age. At the histological examination, fibre size variability was present. The main feature was a severe reduction of the histochemical reaction for both COX and SDH, not associated with ragged red fibres. The biochemical examination showed severe decrease of all the MRC complexes (I, II, III, IV), with strong increase in citrate synthase (CS) activity (online supplementary figure S1A). A second muscle biopsy was performed at 22 years, confirming the histological and histochemical findings (figure 1A-C). At this age, the reduction in the MRC complex activities was still present, yet normal CS activity was observed (online supplementary figure S1B). Furthermore, the activity of PDHC was reduced (PDH/CS: 1.8, normal values: 2.5–5.0). After the genetic identification of the ISCU variant (see below), we carried out histological analysis for the presence of iron deposits in the muscle biopsy by Perls staining. We found Perls-positive material in numerous patient’s fibres, whereas no such material was detected in control muscle biopsies, indicating iron overload in ISCU-mutant muscle (figure 1D).
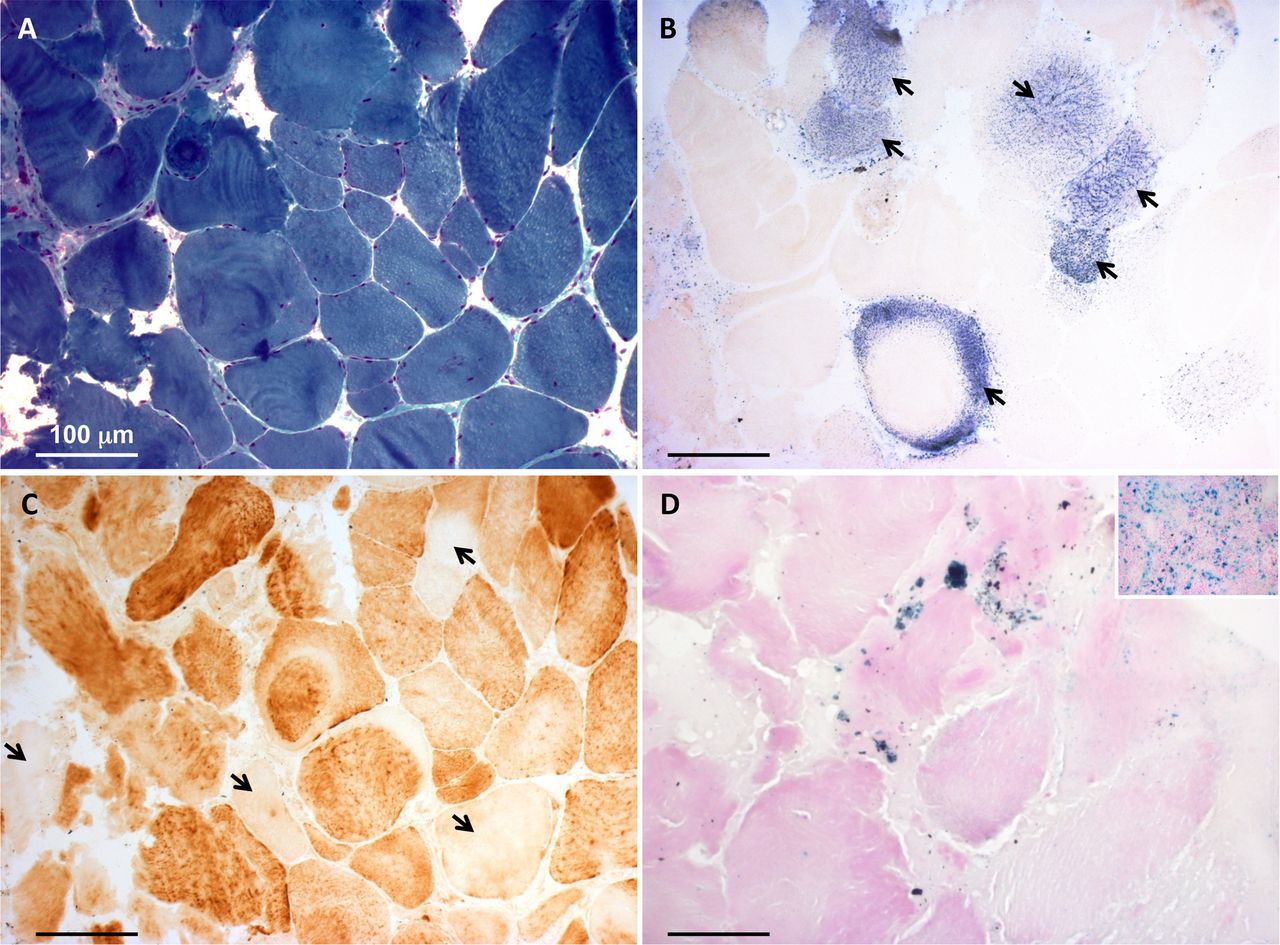
Histochemical analysis of the patient’s muscle biopsy. (A) Gomori trichrome stain showing fibre size variability. (B) Strongly reduced histochemical activity of succinate dehydrogenase. Few fibres showed succinate dehydrogenase-positive staining (arrows). (C) Cytochrome c oxidase (COX) staining showing scattered fibres with severe reduction of histochemical COX activity (arrows). (D) Perls staining demonstrating punctuate accumulation of iron in the patient’s muscle fibres. Inset is a positive control (spleen) for the Prussian blue reaction. Bars correspond to 100 µm.
Genetic studies
Genetic alterations linked to mitochondrial DNA were ruled out: no mutation was detected by sequencing and no evidence of depletion or deletion was observed by Southern blot analysis of mitochondrial DNA from muscle. We performed WES on genomic DNA from the proband. After filtering steps to exclude common SNPs (frequency >0.5%), we selected genes with two compound heterozygous or one homozygous variant, according to a predicted recessive mode of inheritance. Then we focused on genes encoding proteins with mitochondrial localisation. Two entries were found: MTIF2 (translation initiation factor IF-2, mitochondrial) and PDPR (pyruvate dehydrogenase phosphatase regulatory subunit). However, the two missense variants in MTIF2 were in cis, on the same paternal allele, whereas the two variants in PDPR were not confirmed by Sanger sequencing, being probably due to the presence of a pseudogene. No hemizygous variant, suggestive of an X-linked transmission, was detected. A deep analysis of the heterozygous variants, prioritising genes associated with mitochondrial myopathies, highlighted a single variant in ISCU, a c.287G>T (NM_213595.2) predicted to cause the amino acid substitution p.G96V (figure 2A). This nucleotide change was not reported in public variant databases (dbSNP, EVS, ExAC (August 2016)); the substitution affected a highly conserved residue (figure 2B) and gave high scores of pathogenicity, according to several bioinformatics tools. This variant was confirmed by Sanger sequencing in the proband but was not present in the parents’ blood DNA, indicating a de novo event (figure 2C, online supplementary figure S2): the parental DNA samples were analysed also by deep sequencing to exclude very low level of the variant, suggestive of germinal mosaicism. All the ISCU coding regions were well covered by WES. We then screened our patient and his parents for the intronic region encompassing the common mutation present in all the previously described ISCU mutant patients but no variant was identified.

Identification and characterisation of an ISCU mutation. (A) Schematic representation of the ISCU cDNA (NM_213595.2) and ISCU protein with the nucleotide/amino acid change identified in this study. The functional IscU-like domain is in red; the mitochondrial targeting sequence (MTS) is in yellow. (B) Phylogenetic conservation of the amino acid residue (Gly96, in green) affected by the missense mutation identified in the patient. (C) Electropherograms of the genomic region (gDNA) and transcript (cDNA) harbouring the ISCU mutation, and pedigree. (D) Immunoblot analysis of ISCUG96V mutant protein expression and subcellular localisation. Control and patient fibroblasts were harvested by trypsination, permeabilised by digitonin treatment, and separated into a cytosolic and a mitochondria-containing membrane fraction. Total cell lysates as well as cytosolic and crude mitochondrial fractions were subjected to sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE) and analysed for ISCU and ATP synthase F1β subunit (mitochondrial marker) steady-state protein levels (left panel). Chemiluminescence signals of ISCU and F1β in total lysate samples were quantified, and values obtained from patient fibroblasts were expressed relative to control cells (right panel). Error bars indicate the SDs (n=3). UTR, untranslated region.
To exclude that WES could have missed the presence of another deep intronic variant affecting the splicing or a ‘non-exonic’ variant impairing mRNA transcription/stability, we further investigated patient’s specimen at the transcriptional level. No aberrant mRNA ISCU species was observed in PCR products obtained from fibroblast RNA, and their sequencing showed a biallelic expression, suggested by the presence of overlapping peaks corresponding to G and T nucleotides in position c.287 (online supplementary figure S2).
Characterisation of patient’s fibroblasts
The amount of ISCU protein in patient’s fibroblasts was similar to controls, indicating that the mutant protein is normally synthesised, imported into mitochondria and stable (figure 2D). In line with previous reports showing that the biochemical phenotype associated with mutant ISCU is very much attenuated, fibroblasts harbouring the ISCU G96V mutation showed hardly any defect in activities or protein amounts for a number of mitochondrial Fe-S dependent enzymes including mACO, LIAS (as indicated by the presence of lipoate cofactor (Lip) on pyruvate and ketoglutarate dehydrogenase E2 subunits), ferrochelatase and respiratory chain complexes I, II and III (online supplementary figure S3A–F). A minor defect was observed in complex IV, in line with the muscle biopsy analyses, and in the activity of cytosolic aconitase (IRP1). Analysis of steady-state levels of cytosolic/nuclear Fe-S proteins as a measure for maturation-dependent stability revealed no general alteration of their cellular abundance (online supplementary figure S3G-H). However, we observed a severe deficiency of the base-excision DNA repair enzyme NTHL1, a [4Fe-4S] protein, and a slight decrease in protein levels of the CIA factor IOP1 containing 2 [4Fe-4S] clusters. Taken together, our analyses indicate that the presence of the ISCU G96V mutation does not have a strong impact on Fe-S cluster assembly in patient’s cultured fibroblasts.
Yeast model
To assess the pathogenic role of the substitution p.Gly96Val identified in the patient, we performed studies in a yeast model, by introducing the analogous amino acid substitution (G97V) in the Saccharomyces cerevisiae orthologue of ISCU, the yeast gene ISU1. ISU1 has a paralogue, ISU2, arising from a recent gene duplication. The double deletion mutant isu1Δisu2Δ is unviable, thus indicating the essential role of these proteins in the biogenesis of Fe-S clusters, which is in turn indispensable for yeast cell survival.34 The double deletion mutant isu1Δisu2Δ, harbouring the centromeric pFL38 plasmid (URA3 marker) with the wild-type (wt) ISU1 to allow viability, was additionally transformed with pFL39 centromeric plasmids (TRP1 marker) containing either the mutant allele isu1G97V , a wt copy of ISU1, or no gene.
The different strains were plated on 5-fluoroorotic acid containing medium to select for cells that have lost the pFL38/ISU1 plasmid. The strain expressing isu1G97V as the sole ISU1 gene was able to grow on glucose at rates similar to strains carrying the wt ISU1, while the empty pFL39 did not support growth (figure 3A). This result indicates that glycine 97 is not essential for the function of the Isu1 protein. However, growth of the strain expressing the isu1G97V variant was severely retarded on non-fermentable carbon sources (figure 3A), highlighting a deleterious effect of the G97V mutation on mitochondrial function.

Growth analysis of mutant yeast strains. (A) The strain isu1Δisu2Δ harbouring plasmid pFL39 with the wild-type ISU1 gene or the mutant allele isu1 G97V was analysed for growth on various media. Equal amounts of serial dilutions of cells from exponentially grown cultures were spotted onto yeast nitrogen base (YNB) medium plus 2% glucose, 2% lactate or 2% glycerol. The growth was scored after 3 days of incubation at 28°C. (B) The strains isu1Δisu2Δ/ISU1 and isu1Δisu2Δ/isu1 G97V were transformed with pFL38/ISU1 or with the empty vector. Equal amounts of serial dilutions of cells from exponentially grown cultures were analysed for growth on YNB medium plus 2% glucose or 2% lactate after 4 days of incubation at 28°C. (C) Cell yield was calculated by growing cells on liquid medium containing glucose or lactate and measuring the optical density at 600 nm after 72 hours of growth. Error bars indicate the SDs (n=3).
To test whether the G97V mutation acts as a dominant trait, the isu1Δisu2Δ/isu1G97V and the isu1Δisu2Δ/ISU1 strains were transformed with either the pGL38 empty vector (as a control) or pFL38/ISU1 thus obtaining the heteroallelic strain isu1Δisu2Δ/ISU1/isu1G97V and the homoallelic strain isu1Δisu2Δ/ISU1/ISU1. Growth on non-fermentable carbon sources was clearly reduced in the heteroallelic strain compared with the homoallelic wt strain, and also to the strain transformed with the empty vector and expressing a single copy of ISU1 (isu1Δisu2Δ/ISU1). These results indicate that the G97V mutation behaves as dominant (figure 3B). The measurement of the cell yield in liquid cultures confirmed what observed in the spot assay analysis (figure 3C).
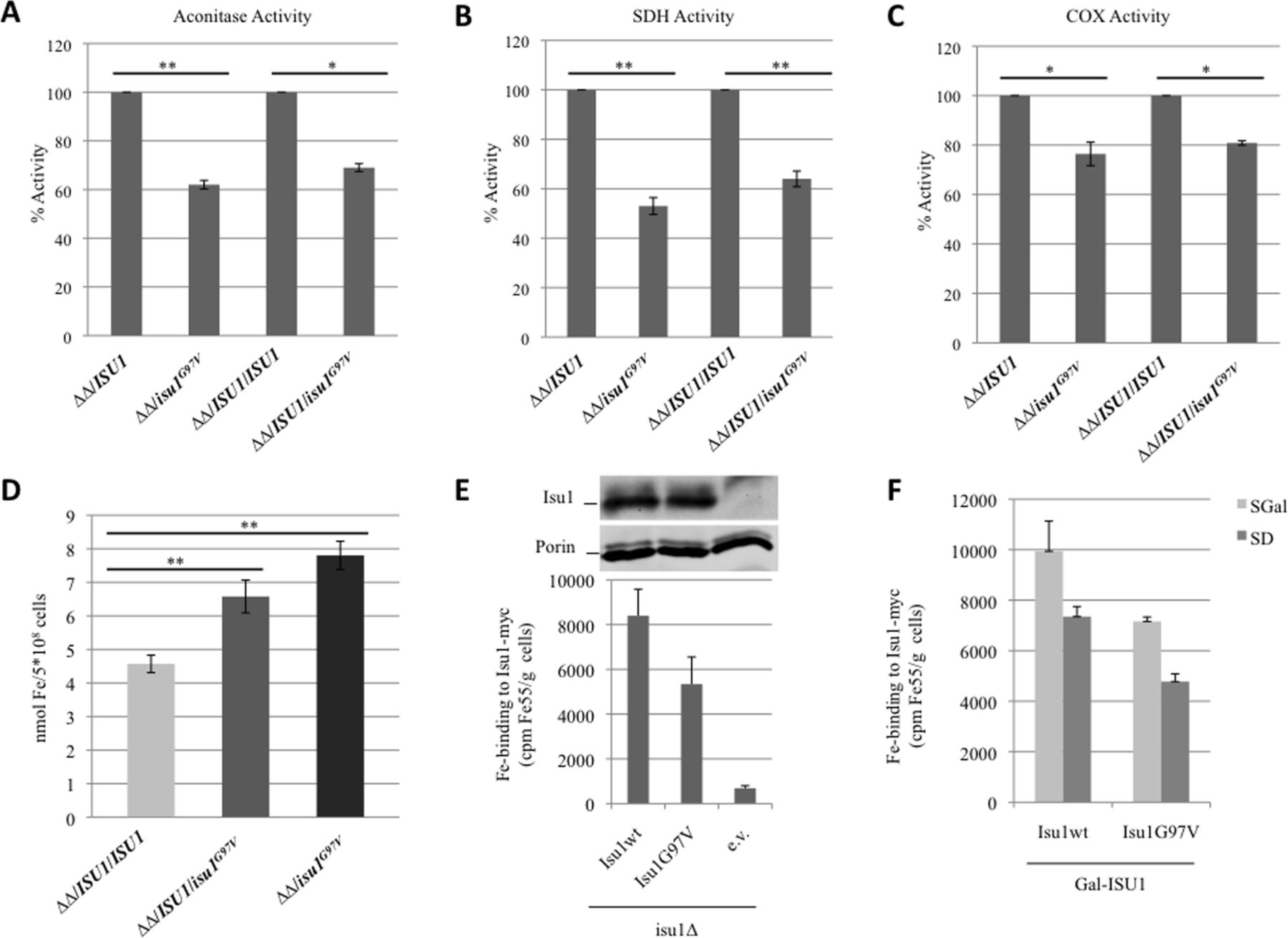
To investigate if the G97V mutation affects mitochondrial Fe-S protein biogenesis, we measured the biochemical activities of two Fe-S cluster-containing enzymes: aconitase and complex II. These activities were reduced in both the isu1Δisu2Δ/isu1G97V and the heteroallelic strain isu1Δisu2Δ/ISU1/isu1G97V (figure 4A and B) indicating an impairment in Fe-S cluster biogenesis in isu1G97V -containing yeast cells. The activity of complex IV which contains two heme centres was also partially reduced (figure 4C). Finally, we evaluated the cellular iron content by a colorimetric assay and found a twofold increase in the mutant compared with wt strain (figure 4D) consistent with a defective core ISC machinery.35

Measurement of enzyme activities and iron content in yeast. (A) Aconitase activity was measured in whole-cell extracts from cells grown exponentially at 28°C in yeast nitrogen base (YNB) medium plus 0.6% glucose. (B and C) Succinate dehydrogenase activity and cytocrome c oxidase activities were measured in a mitochondria-enriched fraction obtained from cells grown as described before. The values for isu1Δisu2Δ/isu1 G97V and isu1Δisu2Δ/ISU1/isu1 G97V strains are expressed as percentage of the activities obtained in the strains isu1Δisu2Δ/ISU1 and isu1Δisu2Δ/ISU1/ISU1. (D) Cellular iron content was quantified in cells grown up to early stationary phase in YNB 0.2% glucose and 2% galactose medium. *<0.05 (unpaired two-tailed t-test), **<0.01 (unpaired two-tailed t-test). (E) Gal-ISU1/isu2Δ cells and isu1Δ cells expressing Myc-tagged Isu1 were radiolabelled with 55Fe and 55Fe incorporation into Isu1-Myc was determined by immunoprecipitation with α-Myc antibodies followed by scintillation counting. Wild-type cells harbouring the empty vector (e.v.) served as control. Isu1-myc protein levels in isu1Δ cells were determined by immunostaining with α-Myc antibodies. Porin (Por1) served as a loading control. (F) Gal-ISU1/isu2Δ cells expressing Isu1 from vector pFL39 and the reporter plasmid pFET3-GFP were cultivated in SD or SGal medium supplemented with 50 µM ferric ammonium citrate. At an optical density=0.5, the GFP-specific fluorescence emission of whole cells was determined. Error bars indicate the SDs (n=3).
To further investigate the impact of the G97V mutation on Isu1 function, we measured the ability of mutant versus wt Isu1 protein to assemble a Fe-S cluster in vivo.36 To this end, we used the strain Gal-ISU1/isu2Δ, in which ISU2 is deleted and ISU1 is under the control of the GAL1-10 promoter. The levels of Isu1 can be downregulated by growing the cells in the presence of glucose.37 Gal-ISU1/isu2Δ cells were transformed with pRS426-TDH3 vectors encoding Myc-tagged ISU1wt or isu1G97V . To estimate Fe-S cluster binding of Isu1, cells were radiolabelled with 55Fe, and the incorporation of radioactivity into Isu1G97V or Isu1wt was measured by immunoprecipitation and scintillation counting (figure 4E,F). As shown in figure 4F (darker bars), the mutant protein showed a 30% reduction in the 55Fe binding capacity; this reduction is consistent with the mutant growth defect displayed on respiratory carbon sources. A decrease was also evident when isu1G97V was expressed in isu1Δ cells or when Gal-ISU1/isu2Δ cells were depleted by growth in galactose-containing medium, conditions in which endogenous wt Isu1 and Isu2 are present, thus mimicking the heteroallelic condition (figure 4F, light grey bars, and figure 4E). This result showed that the Isu1G97V mutant protein is slightly impaired in its ability to incorporate Fe into Fe-S clusters in vivo. The alterations observed in mutant strains were not due to a reduced amount of the mutant Isu1G97V protein since its abundance was similar to the wt form (figure 4E). Furthermore, Gal-ISU1/isu2Δ cells expressing isu1G97V from vector pFL39 displayed a twofold increase of the iron-dependent FET3 promoter on both glucose and galactose-containing medium (online supplementary figure S4). A deregulated iron homeostasis is a hallmark of cells with defective core ISC machinery and explains the increased iron levels in cells expressing isu1G97V .
In silico pathogenicity prediction and structural modelling
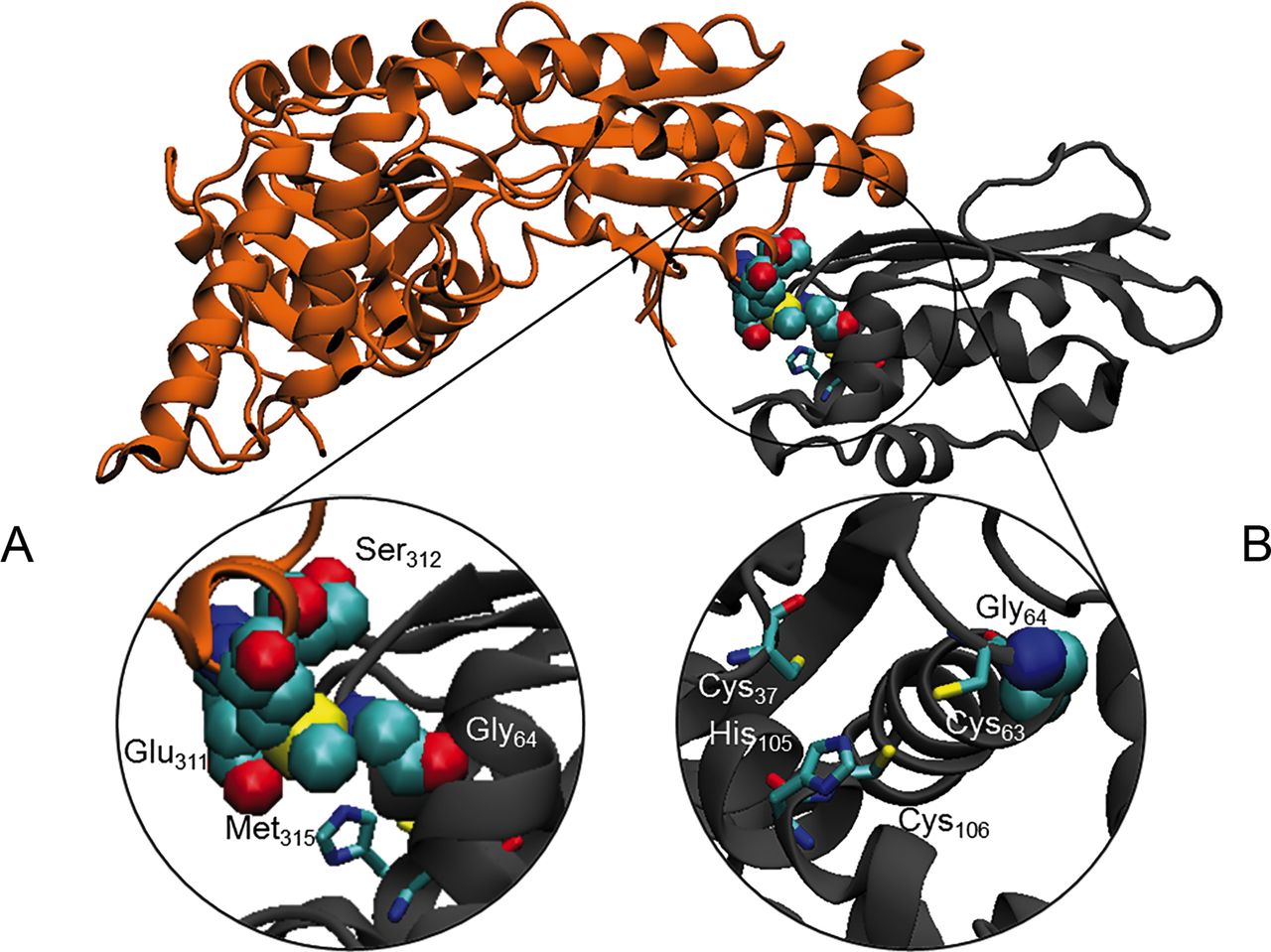
In silico predictions, by using different tools, were used to evaluate the potential effect of the Gly96Val substitution on ISCU structure and interaction with its protein partners. ISCU interacts with NFS1, yet the 3D structure of the human NFS1-ISCU complex is hitherto unknown. Instead, we used the reported crystal structure of the E. coli IscS-IscU complex formed by an IscS dimer which binds two IscU at opposite ends of IscS.38 Notably a similar, yet unusual monomeric structure has been recently described for the IscS human orthologue NFS1.39 Human Gly96 which corresponds to Gly64 in E. coli 38 is located in a flexible loop, at the beginning of the helix α1, next to the potential Fe-S cluster-co-ordinating Cys95 residue (Cys63 in E. coli).40 We analysed the possible interactions of Gly64 with IscS/NFS1 by using First Glance in Jmol. The glycine backbone could establish hydrophobic interaction with IscS Met315 that is located inside a stretch (YVEGESLLMAL) highly conserved from E. coli to human. Although both glycine and valine are hydrophobic residues, the greater steric hindrance of the valine side chain could interfere with the IscS-IscU interaction (figure 5A). Moreover, the loop encompassing residues 62–64 (GCG) of IscU is opposite to Glu311 and Ser312 on IscS, and we suggest that the valine slightly disturbs these hydrophilic residues (figure 5A). The substitution Gly96Val could therefore prevent the correct orientation of the adjacent cysteine side chain, thus interfering with the Fe co-ordination and explaining the defective ISCU function (figure 5B).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
In silico structural analysis. (A) Ribbon diagram of the Escherichia coli IscS (coloured in grey)-IscU (coloured in orange) complex (PDB ID: 3lvl). Close view of the Gly64 of IscU and Glu311, Ser312 and Met315 of IscS represented in Van der Waals and coloured by type. (B) Residues supposed to be involved in the co-ordination of the 2Fe-2S are represented as sticks.
Discussion
ISCU (Isu1-2 in yeast) is an essential scaffold protein for the biosynthesis of Fe-S clusters. Recessive hypomorphic ISCU alleles have been associated with isolated myopathy11 12 or skeletal and cardiac myopathy14 in humans. Here, we report a heterozygous, de novo dominant ISCU variant, G96V, causing congenital myopathy in a single patient. The following considerations provide evidence that this variant exerts a deleterious, dominant effect on Fe-S-dependent enzyme activities. First, we observed striking clinical, histological, histochemical and biochemical abnormalities affecting skeletal muscle, which define a mitochondrial myopathy similar to that reported in ISCU recessive mutations, including partial depletion of SDH and COX histochemical reactions, generalised reduction of the MRC complex activities, and accumulation of iron deposits. Similar to recessive ISCU cases, our patient displayed isolated myopathy with fluctuating, waxing and weaning episodes of profound muscle weakness, in the context of a congenital myopathy with moderately high CK and no involvement of extramuscular organs, including the CNS. Second, the mutation was absent in both healthy parents, supporting its sporadic occurrence in the proband, and was the only gene defect related to mitochondrial myopathies identified by WES. Third, analysis of the corresponding genetic defect in yeast suggested a Fe-S protein biogenesis defect, including defects in OXPHOS and the cellular iron regulon. The effects were dominant, since both the monoallelic and the heteroallelic genotypes were associated with the phenotype.
While our yeast studies suggest that Isu1G97V is functionally impaired by itself, what might then be the reason for the observed dominance of both the human ISCUG96V and corresponding yeast Isu1G97V mutations? ISCU, as the scaffold protein for the de novo synthesis of Fe-S clusters within mitochondria, interacts with a number of other ISC proteins during Fe-S biosynthesis, although the stoichiometry of these interactions is controversial. Similar to the bacterial structure, the mammalian ISC complex was proposed to be composed of two NFS1 and two ISCU subunits,4 41 with a central NFS1 dimer and two molecules of ISCU at each end of the NFS1 dimer. Recently, a [FXN42–210]24-[NFS1]24-[ISD11]24-(ISCU)24 complex model was proposed,40 but it was obtained by overexpressing human proteins in E. coli and it is probably not relevant in vivo. We think that the functional impairment of ISCU by the G96V mutation may be caused by structural changes, as suggested by our in silico analyses. For instance, the orientation of the potential Fe-S cluster-co-ordinating Cys95 adjacent to Gly96 could be altered by replacement of the helix-breaking glycine residue, with a negative outcome on protein function in Fe-S cluster assembly. Moreover, the mutated protein structure may become stiffer, thus negatively affecting the interactions with its other partner ISC proteins, notably NFS1. The altered interaction between NFS1 and the mutant ISCU could in turn affect the assembly/function of the entire ISC biosynthetic complex. In support of this view stands the observation that the ISCUG96V mutant protein is stable and not degraded, and its amount is similar to that of ISCUwt in patient’s fibroblasts. In addition, the dominant effect may be caused by the intermediate formation of an ISCU holodimer which, according to in vitro reconstitution studies, is the product of the ISC biosynthetic complex.5 42 In the homodimer, the mutated Gly96 residue in one ISCU monomer would likely directly face the other wt ISCU monomer because of the bridging character of [2Fe-2S] cluster binding. This could lead to the observed dominant negative effect of the mutated ISCU on wt ISCU proteins. This might also be the reason why we observed less radiolabelled 55Fe bound to yeast Isu1G97Vcompared with the wt protein. Interestingly, the dominant-negative behaviour of the G97V mutation differs from the recessive effect reported for the G50E mutation in both yeast and humans, which can be ascribed to haploinsufficiency of the G50E allele.43
Although ubiquitous, the ISCUG96V variant produces a clinically detectable effect only in one critical tissue, skeletal muscle, in line with the purely myopathic presentation of the common ISCUIVS5+382G>C splicing variant already reported in Swedish patients. Similar to our results, no evident mitochondrial phenotype in fibroblasts has been previously reported in cases with ISCU-related recessive disease.14 44–46 The molecular mechanisms that make skeletal muscle exquisitely sensitive to partial ISCU impairment warrant further investigation in patient-derived cells and, possibly, animal models. A tissue-specific splicing of ISCU was proposed for the common ISCUIVS5+382G>C mutation to explain the skeletal muscle phenotype,13 but this hypothesis cannot be applied for either the p.G96V or the p.G50E missense variants, which are equally associated with muscle-specific dysfunction. The structural alterations and impaired interactions with partner proteins caused by the p.G96V could have tissue-specific effects which explain the muscle involvement. A greater sensitivity of ISCU-mutant myoblasts to oxidative stress,45 and exercise-induced oxidative damages in muscle,47 may also account for the muscular phenotype. Nevertheless, tissue specificities for many of the mitochondrial diseases due to defective nuclear-encoded genes with housekeeping functions are currently poorly understood and hardly predictable.
The multiple biochemical defects of MRC activities observed in our patient were expected for complexes I, II and III which contain Fe-S clusters. A defect was also seen for the Fe-S cluster-free complex IV, which harbours Cu2+ and Fe-containing heme a/a3 as redox centres. In principle, this may be due to a defect in the Fe-S protein ferrochelatase catalysing the last step of heme biosynthesis.48 However, strong complex IV defects are also seen on depletion of ISCA1, ISCA2 and IBA57 in both human and yeast cells.49 50 These ISC proteins are specific for mitochondrial [4Fe-4S] protein maturation, yet are not involved in the assembly of the [2Fe-2S] cluster on human ferrochelatase. Moreover, we observed a complex IV impairment in the mutant yeast model despite the yeast orthologue enzyme is devoid of a Fe-S cluster. Nevertheless, complex IV defects are common secondary effects of an impairment of Fe-S protein biogenesis.6 49 A complex IV deficiency (19%–28% residual activity) was also reported in the two siblings with the most severe ISCU-related phenotype, and a partial reduction was found in five subjects with the Swedish-type myopathy.14 This may be due to a downstream damaging effect linked to impairment of complexes I, II and III or to the hampered formation of MRC supercomplexes. In addition to the biochemical defects, the histochemical analysis was peculiar, with a quite specific pattern of SDH and COX deficiency; this was suggested to be a pathognomonic finding of a myopathy related to Fe-S cluster.9 However, in patients with mutations in FXN, the picture is different, with mainly COX negative fibres and nearly normal SDH staining.51 Other Fe-S diseases are rare and usually present as neurological disorders with minimal myopathic signs, while little is known about their muscle features. Collectively, the histochemical pattern in muscle biopsy seems specific for ISCU myopathy.
In conclusion, we report the first heterozygous dominant mutation in ISCU; notably, this alteration resulted in a similar phenotype as the recessive ISCU disease previously described. Our finding stresses the importance of a deep analysis of WES data that may include, for sporadic cases, any mode of transmission. Moreover, our study confirms that recessive and dominant mutations in the same gene may lead to the same disease, as already reported for other mitochondrial disorders (eg, DNM1L mutations).
Acknowledgments
We are grateful to Ileana Ferrero for stimulating discussions, Joanna Poulton for providing patient’s fibroblasts, the contribution of the Core Facility ‘Protein Spectroscopy and Protein Biochemistry’ of Philipps-Universität Marburg, and R. Rösser for excellent technical support. We acknowledge the ‘Cell lines and DNA Bank of Paediatric Movement Disorders and Neurodegenerative Diseases’ of the Telethon Network of Genetic Biobanks (grant GTB12001J) and the Eurobiobank Network for supplying biological specimens.
References
Footnotes
AL and AR contributed equally.
Contributors AL carried out the molecular biology experiments; AR carried out WES; CCB performed yeast studies under the supervision of PG; OS performed experiments in patient’s fibroblasts; SM carried out the histochemical analysis; CL provided the clinical report; AF performed structural analysis; AJR provided bioinformatics management of the WES data; UM performed yeast studies under the supervision of RL; RL, MZ, PG and DG organized the experimental set-up and wrote the manuscript; all authors read and approved the manuscript.
Funding This work was supported by the TelethonItaly [GrantGGP15041]; the Pierfranco and Luisa Mariani Foundation; the MRC7QQR [201572020] grant; the ERC advanced grant [FP77322424]; the NRJ Foundation7Institut de France; the E7Rare project GENOMIT. RL acknowledges generous financial support from Deutsche Forschungsgemeinschaft [SFB 987 and SPP 1927] and the LOEWE program of state Hessen.
Competing interests None declared.
Patient consent Obtained.
Ethics approval Ethical Committee of the Fondazione IRCCS Istituto Neurologico ‘Carlo Besta’, Milan, Italy.
Provenance and peer review Not commissioned; externally peer reviewed.
Data sharing statement Values of the biochemical activities reported in the graphs are available upon request.Images of the histochemical stainings are available upon request. The complete list of the variants found by WES in the proband is available upon request.
Correction notice This article has been corrected since it was published Online First. The affiliation of Massimo Zeviani has been updated.