Article Text

Abstract
Background The interpretation of germline TP53 variants is critical to ensure appropriate medical management of patients with cancer and follow-up of variant carriers. This interpretation remains complex and is becoming a growing challenge considering the exponential increase in TP53 tests. We developed a functional assay directly performed on patients’ blood.
Methods Peripheral blood mononuclear cells were cultured, activated, exposed to doxorubicin and the p53-mediated transcriptional response was quantified using reverse transcription–multiplex ligation probe amplification and RT-QMPSF assays, including 10 p53 targets selected from transcriptome analysis, and two amplicons to measure p53 mRNA levels. We applied this blood functional assay to 77 patients addressed for TP53 analysis.
Results In 51 wild-type TP53 individuals, the mean p53 functionality score was 12.7 (range 7.5–22.8). Among eight individuals harbouring likely pathogenic or pathogenic variants, the scores were reduced (mean 4.8, range 3.1–7.1), and p53 mRNA levels were reduced in patients harbouring truncating variants. We tested 14 rare unclassified variants (p.(Pro72His), p.(Gly105Asp), p.(Arg110His), p.(Phe134Leu), p.(Arg158Cys), p.(Pro191Arg), p.(Pro278Arg), p.(Arg283Cys), p.(Leu348Ser), p.(Asp352Tyr), p.(Gly108_Phe109delinsVal), p.(Asn131del), p.(Leu265del), c.-117G>T) and 12 yielded functionally abnormal scores. Remarkably, the assay revealed that the c.*1175A>C polymorphic variant within TP53 poly-adenylation site can impact p53 function with the same magnitude as a null variant, when present on both alleles, and may act as a modifying factor in pathogenic variant carriers.
Conclusion This blood p53 assay should therefore be a useful tool for the rapid clinical classification of germline TP53 variants and detection of non-coding functional variants.
- germ-line mutation
- genetic testing
- genetic predisposition to disease
- methods
- clinical laboratory techniques
Data availability statement
All data relevant to the study are included in the article or uploaded as supplementary information. Deidentified participant data are available from thierry.frebourg@chu-rouen.fr.
This is an open access article distributed in accordance with the Creative Commons Attribution Non Commercial (CC BY-NC 4.0) license, which permits others to distribute, remix, adapt, build upon this work non-commercially, and license their derivative works on different terms, provided the original work is properly cited, appropriate credit is given, any changes made indicated, and the use is non-commercial. See: http://creativecommons.org/licenses/by-nc/4.0/.
Statistics from Altmetric.com
- germ-line mutation
- genetic testing
- genetic predisposition to disease
- methods
- clinical laboratory techniques
Introduction
Identification of a germline pathogenic TP53 (MIM: *191170) variant in a patient with cancer has drastic medical impacts.1 Indeed, in TP53 variant carriers, chemotherapy and radiotherapy have been shown to contribute to the development of subsequent primary cancers, the incidence of which is remarkably high (above 40%).1–4 Therefore, in these patients, surgical treatment should be prioritised and radiotherapy and chemotherapy avoided, if possible, or at least carefully discussed in terms of benefit:risk ratio between risk of recurrence and risk of inducing second primary tumours. Furthermore, TP53 variant carriers should have specific surveillance protocols, including annual whole-body MRI,5 6 whose efficiency for early tumour detection has recently been shown by numerous studies.5–14
Interpretation of germline TP53 variants, which are mainly missense variants, remains particularly complex. Whereas germline variants of TP53 were initially detected in Li-Fraumeni syndrome (LFS, MIM#151623),15–17 our perception of cancers related to germline alterations of TP53 has drastically evolved over time.1 2 18 19 The presence of a disease-causing germline variant should be considered in patients fulfilling Chompret criteria, which were sequentially updated and extended.1 The question of germline TP53 variant interpretation is becoming a growing concern in the field because the TP53 gene is currently included in many cancer gene panels, and the number of TP53 tests performed in patients not fulfilling the criteria mentioned earlier has increased exponentially. 20 21
Classification of TP53 variants, in agreement with the American College of Medical Genetics and Genomics/Association for Molecular Pathology guidelines, is based on several items, including frequency of the variant in the general population (gnomAD; https://gnomad.broadinstitute.org/), segregation data, bioinformatics predictions and functional assays developed in yeast or human cancer cell lines.22 One of the first assays commonly used for TP53 missense variant interpretation was developed in yeast and is based on the expression of TP53 cDNA in strains containing reporter plasmids with different p53 binding sites.23 In this assay, p53 variants are classified as functional, not functional or partially functional if the transcriptional activity is conserved for some but not all yeast reporter plasmids (http://p53.iarc.fr/). More recently, two teams have developed in human cancer cell lines high throughput p53 functional assays.24 25 Kotler et al 24 generated a synthetic library of TP53 variants located within the p53 DNA-binding domain and quantified the antiproliferative activity of these variants in the p53-null H1299 cancer cell line. In this assay, TP53 variants are categorised as ‘wild-type TP53-like variant’ (functional) or ‘disrupting’ (non-functional). In another assay, Giacomelli et al 25 generated by saturation mutagenesis a TP53 library and tested the ability of the variants (1) to restore the survival of the p53-null A459 cell line exposed to high doses of DNA damaging agents, in order to detect loss of function (LOF) variants and (2) to induce in p53-wild-type A459 cells resistance to Nutlin-3, in order to detect variants with dominant negative effect (DNE).
We previously developed, in Epstein-Barr virus-immortalised lymphocytes, a p53 functional assay exploring the transcriptional activity of the protein underlying its tumour suppressor activity.26 This assay is based on the exposure of cells to DNA damaging agents followed by the measurement of the p53 transcriptional response.27 28 With this assay, we showed that pathogenic TP53 variant carriers exhibit a constitutive defect in the transcriptional response to DNA damage, establishing a biological endophenotype associated with germline pathogenic variants.27 28 Compared with the other assays, its main advantage is to evaluate the impact of heterozygous variants in the genetic context of the patients. Its main disadvantage is that it requires EBV immortalisation, which is time-consuming and, therefore, not suited for a rapid classification and interpretation of TP53 variants in medical practice.
Therefore, despite the different tools indicated previously and before the completion in the future of curated international databases, interpretation of germline TP53 variants remains challenging in clinical practice. This prompted us to develop a p53 functional assay derived from the previous one but performed on fresh blood samples and suitable for rapid interpretation and medical management of patients. We show here that this assay can accurately detect pathogenic variants and can be used to reallocate unclassified variants by integrating the results to the classification strategy.22 Furthermore, this assay revealed that a TP53 polymorphism (rs78378222), present in 1.7% of the European population, compromises p53 functional activity with the same magnitude as a heterozygous null variant, when carried on both alleles.
Methods
Cell culture and treatment
EBV-immortalised cell lines were maintained in RPMI 1640 medium (GIBCO; Life Technologies, Carlsbad, California, USA) with 10% fetal calf serum (Invitrogen, Life Technologies) and 1% L-glutamine (Invitrogen) at 37°C with 5% CO2. Cells were seeded in duplicate in 12-well plates (Corning, New York, USA) at a density of 106 cells/well. Cells were treated or not with 200 ng/mL (0.3 µM final concentration) of doxorubicin (Sigma Aldrich, St. Louis, Missouri, USA) for 8 hours. Cells were washed with 1× PBS and harvested for RNA extraction.
Peripheral blood mononuclear cell (PBMC) isolation and culture
Blood samples were collected in EDTA tubes and kept for 2 days at room temperature before PBMC isolation on a lymphocyte separation medium (Eurobio, Evry, France). From 2.5 to 10.0 mL of blood per patient was used for PBMC isolation. Cell number and cell viability were assessed on a NanoEnTek Adam automatic cell counter with the AccuChip Kit (ScienceTEC, Villebon-sur-Yvette, France). One million cells were seeded per well in a 24-well plate and were let to grow for 48 hours in a lymphocyte activating medium (Chromosome Medium P, AmpliTech, Compiègne, France). At least two wells were seeded per patient (treated and untreated) and duplicates or triplicates were performed whenever possible. Cells were treated with 800 ng/mL of doxorubicin for 8 hours, washed with 1× PBS, harvested and RNA extraction was performed using the NucleoSpin RNA XS kit (Macherey Nagel, Düren, Germany) according to the manufacturer’s instructions and quantified using a UV-VIS ND-1000 spectrophotometer (Biocompare, Nanodrop Technologies, USA).
RNA-Seq
Four control EBV cell lines wild-type for TP53 and four heterozygous TP53-mutant cell lines, corresponding to three canonical dominant negative missense variants (p.(Arg175His), p.(Arg248Trp) and p.(Arg273His)) and one complete deletion of the TP53 locus, were treated or not with doxorubicin. RNA was extracted using the Nucleospin RNAII kit (Macherey Nagel). Libraries were prepared using the NEBNext Ultra Directional RNA Library Kit for Illumina (NEB, Ipswich, USA) and NGS sequencing of the libraries was performed on an Illumina NextSeq500 (Illumina, San Diego, USA) using 2*75 bp sequencing to generate 50M read pairs on average per sample. Experiments were performed in triplicates. Bioinformatic analysis was carried out using an in-house automated pipeline AURIGA that uses the STAR V.2.5.3a tool for alignment, FeatureCounts tool V.1.5.2 for read counting and DESeq2 V.1.18.1 for statistical analysis.
Selection of biomarkers indicative of p53-transcriptional activity
New biomarkers were selected among the transcripts strongly up-egulated by doxorubicin in control cells but not in the cells harbouring heterozygous TP53 alterations: CEP170B (NM_015005), PODXL (MIM*602632, NM_001018111), RRAD (MIM*179503, NM_004165), GLS2 (MIM*606365, NM_013267), CABYR (MIM*612135, NM_012189), TP53I3 (MIM*605171, NM_004881), EPS8L2 (MIM*614988, NM_022772), SULF2 (MIM*610013, NM_001161841), SESN1 (MIM*606103, NM_014454) and FHL2 (MIM*602633, NM_201555). Three control transcripts with a steady expression across all conditions and genotypes and expressed at the same level as the selected targets were also selected: TBP (MIM*600075, NM_003194), RIC8B (MIM*609147, NM_001330145) and MPP5 (MIM*606958, NM_022474.3). An internal control of treatment efficacy was included: PLK1 (MIM*602098, NM_005030.5), whose transcript is downregulated by doxorubicin treatment both in wild-type and mutant cells.
Reverse transcription–quantitative multiplex PCR of short fluorescent fragment (RT-QMPSF)
Reverse transcription (RT) was performed on 100 ng of total RNA using the Verso cDNA Synthesis Kit (Thermo Scientific, Waltham, USA). RT-QMPSF was performed on 1.5 µL of RT using Diamond Taq DNA polymerase (Kaneka Eurogentec, Seraing, Belgium), 6% Dymethyl sulfoxide and 26 PCR cycles (94°C: 30 s/58°C: 1 min/72°C: 30 s). Primer sequences are listed in online supplemental table 1. Amplicons were analysed on an ABI Prism 3500 Genetic Analyzer (Applied Biosystems, Foster City, California, USA) using GeneScan 3.7 software.
Supplemental material
Reverse transcription–multiplex ligation probe amplification (RT-MLPA)
RT-MLPA probes were pooled at a concentration of 1 fmol/µL each in 10 mM Tris/1 mM EDTA. Probe sequences are given in online supplemental table 1. RT (6.5 µL), probe mixture (1.5 µL) and SALSA-MLPA buffer (1.5 µL, MRC-Holland, Amsterdam, The Netherlands) were mixed before denaturation (95°C, 2 min) and hybridisation (60°C, 1 hour). Ligation was performed at 54°C for 15 min, adding 32 µL of ligation mixture, and heated 5 min at 98°C. Then, 2.5 µL of the ligation was added to 7.5 µL of a Q5Hot Start High-Fidelity 2X Master Mix (NEB) supplemented with universal fluorescent PCR primers. PCR was performed using 35 cycles (94°C: 30 s/58°C: 30 s/72°C: 30 s). Amplicons were analysed on an ABI Prism 3500 Genetic Analyzer using GeneScan V.3.7 software.
Calculation of p53 functionality score and p53 mRNA ratio
The RT-MLPA or RT-QMPSF profiles of doxorubicin-treated and untreated cells were superimposed after adjusting the control amplicons to the same height. In the treated condition, the peak height of each of the 10 p53 target genes was measured and divided by the sum of the heights of the three control genes. This value was then divided by the same ratio calculated in the untreated condition. In the assay, the mean of the 10 values defines the p53 functionality score. The final p53 functionality score is the mean of the scores obtained in RT-MLPA and RT-QMPSF assays. The p53 mRNA levels were expressed as a ratio of the normal values obtained for 3 TP53 wild-type control individuals. The efficacy of the genotoxic treatment was assessed by calculating a PLK1 (MIM*602098) ratio (treated/untreated) normalised with the three controls, which should be less than 0.5.
Results
Development of a rapid p53 functional assay performed on blood
The rationale of the assay is that p53 acts as a powerful transcriptional inductor when DNA damage occurs and that the common deleterious impact of pathogenic variants is the alteration of this transcriptional activity.26 To develop a functional assay directly performed on patient’s fresh blood, we first optimised the quantitative assay that we had previously developed in EBV-immortalised cell lines.27 28 To this aim, we performed a new comparative transcriptomic analysis using RNA-Seq, including non-polyadenylated RNAs. Four control EBV cell lines wild type for TP53 and four patients with LFS EBV cell lines were compared in the context of genotoxic stress induced by doxorubicin treatment. We selected 10 biomarkers corresponding to p53 targets involved in different biological pathways controlled by p53, such as cell adhesion and migration, cellular response to stress, apoptosis, cytoskeleton organisation, glycolysis or regulation of other metabolic pathways. To normalise the results, we selected three transcripts with a steady expression across all conditions and genotypes. All these biomarkers were then included in two quantitative assays based on RT-MLPA and RT-QMPSF. To detect in the same assay the potential effect of variants on the TP53 transcript levels, we added different amplicons or probes corresponding to TP53 cDNA. As a defect in treatment efficacy would result in a low functionality score leading to the misinterpretation of a wild-type genotype as a mutant one, we also integrated in the assays an internal control of treatment efficacy. After exposure to doxorubicin, cells were harvested and the RT-MLPA and RT-QMPSF assays were performed in parallel for each sample to increase the robustness of the assay. An arbitrary functionality score was calculated from the induction score of the 10 p53 targets; the p53 RNA levels were evaluated and expressed as a percentage of the mean levels obtained for three wild-type TP53 individuals. This new quantitative assay, based on both RT-QMPSF and RT-MLPA, was first validated on 31 lymphoblastoid cell lines derived from patients with LFS harbouring different germline heterozygous TP53 variants (online supplemental table 2).
Supplemental material
We then set up the conditions allowing the assay to be performed directly on the patients’ peripheral blood. Blood was collected in conventional EDTA tubes and kept at room temperature for 2 days to mimic sample shipping delays. PBMCs were isolated and cultured for 48 hours in a lymphocyte activating medium. Under these conditions, a strong p53 transcriptional response could be monitored in wild-type individuals (figure 1), indicating that testing p53 function directly on patients’ blood cells was feasible.

P53 functional assay on peripheral blood. (A) Schematic representation of the blood p53 functional assay workflow. (B,C) Typical RT-QMPSF (B) and RT-MLPA (C) results obtained for individual 15 with a wild-type TP53 genotype. The fluorescent profiles of doxorubicin-treated cells (red line) and untreated cells (blue line) were superimposed using the three control amplicons (RIC8B, TBP and MPP5). The horizontal bars indicate for each p53 target gene the level of expression in untreated cells. Treatment efficacy was evaluated by the transcriptional repression of the PLK1 marker (Plk1 treated/untreated ratio below 0.5). In the treated condition, the peak height of each of the 10 p53 target genes was measured and divided by the sum of the heights of the three control genes. This value was divided by the same ratio calculated in the untreated condition to yield an arbitrary p53 functionality score. The p53 mRNA levels were expressed as a ratio of the normal values obtained for three control individuals. PBMC, peripheral blood mononuclear cell; RT-MLPA, reverse transcription–multiplex ligation probe amplification; RT-QMPSF, reverse transcription–quantitative multiplex PCR of short fluorescent fragment.
p53 functional analysis of patient’s blood cells with different TP53 genotypes
We then applied the p53 functional assay on blood samples sent to our laboratory for TP53 molecular analysis (NGS screening of the 11 exons complemented by QMPSF). Molecular and functional analyses were performed in parallel, in double blind conditions. We analysed a total of 82 blood samples derived from 77 individuals (online supplemental table 3). These 77 individuals corresponded either to new index cases suspected to harbour a pathogenic TP53 variant or to relatives of index cases harbouring TP53 variants. This sample reflects the real-life recruitment of our diagnostic laboratory as it includes unaffected individuals as well as individuals affected by cancer who may have undergone different chemotherapy treatments. Molecular analyses revealed that 51 individuals had no detectable germline TP53 variant. For these 51 individuals, the mean p53 functionality score measured was 12.7 (13.6 for the RT-QMPSF assay and 11.9 for the RT-MLPA assay) with a range of 7.5–22.8 (online supplemental table 3 and figure 2). The mean observed p53 mRNA levels were 93% with a range of 74%–125% (online supplemental table 3). In eight tested individuals, molecular analysis revealed seven distinct TP53 variants which could be considered as likely pathogenic or pathogenic based on their ClinVar classification or their truncating nature (table 1). All the variants tested were confirmed to be germline heterozygous variants. For these eight patients, the assay yielded a reduced score compared with the wild-type individuals (mean 4.8, range 3.1–7.1; table 1 and figure 2). In the patients with missense variants, p53 mRNA levels were above 75%. In contrast, p53 mRNA was clearly reduced in patients harbouring frameshift or splice variants (mean 58%, table 1 and figure 2) probably reflecting the activity of the nonsense-mediated mRNA decay.
Supplemental material
Interpretation of germline TP53 variants integrating the blood p53 functional assay
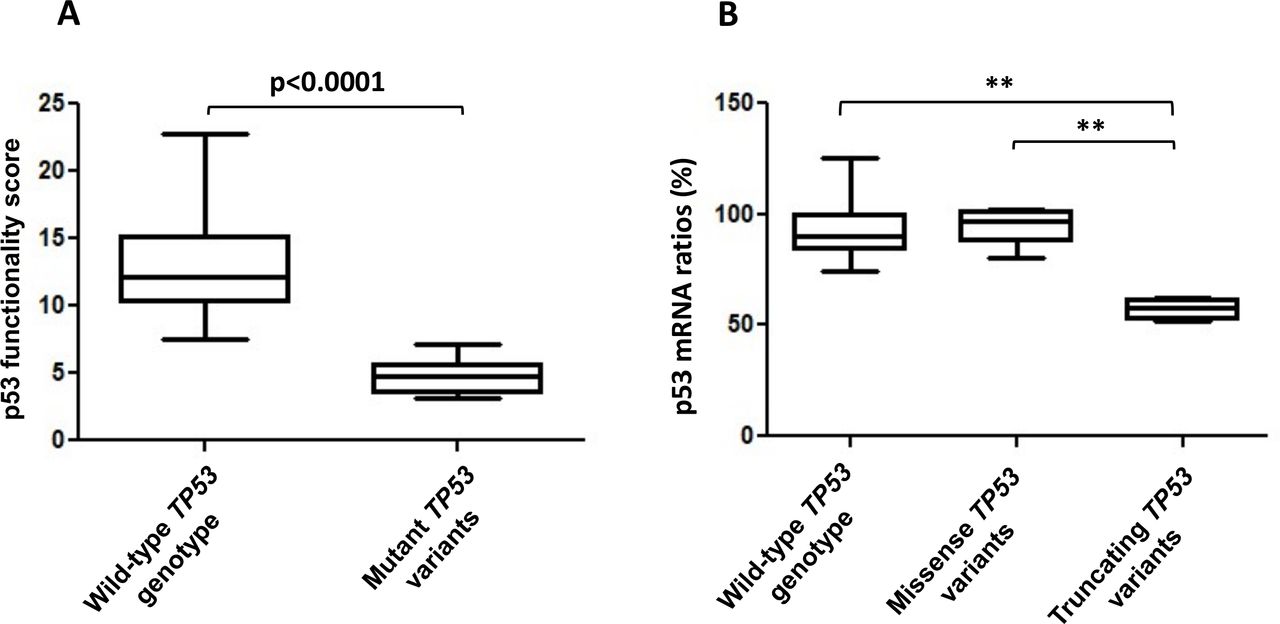
p53 functional scores and mRNA level ratios in individuals with wild-type TP53 or with germline TP53 variants. (A) p53 functionality scores obtained in 51 wild-type TP53 individuals, compared with the scores obtained for nine samples from eight individuals carrying a classified TP53 variant (online supplemental table 3) using a Mann-Whitney non-parametric test. (B) Comparison of the p53 mRNA ratios obtained in 51 wild-type TP53 individuals and in samples carrying a missense (five samples) or a truncating variant of TP53 (four samples), using a Kruskal-Wallis test with Dunns post-test (p=0.0031). ***P<0.01.
Functional evaluation of TP53 variants of unknown biological significance
Based on these results, we refined the experimental thresholds for the analysis of unclassified variants. A functionality score above 7.5 was considered as indicative of a wild-type TP53 genotype, a score below 7.5 indicative of a variant impacting p53 function. A score below 5.5 is associated with a strong impact, and a score between 5.5 and 7.5 is associated with an intermediate effect. A reduction in p53 mRNA levels (<65%) was, by itself, suggestive of a defect independently of the functional score obtained. Using these criteria, we undertook the functional interpretation of 14 rare TP53 variants which remained unclassified despite the available tools (table 1). These variants included 10 rare missense variants (p.(Pro72His), p.(Gly105Asp), p.(Arg110His), p.(Phe134Leu), p.(Arg158Cys), p.(Pro191Arg), p.(Pro278Arg), p.(Arg283Cys), p.(Leu348Ser), p.(Asp352Tyr)), 2 in-frame deletions (p.(Asn131del), p.(Leu265del)), 1 in-frame deletion–insertion [p.(Gly108_Phe109delinsVal)] and 1 rare variant within the 5′UTR region (c.-117G>T). For these rare variants (table 1), except p.(Pro191Arg) and c.-117G>T, the functional score was below 7.5, classifying these variants as ‘functionally abnormal’, according to the terminology recently recommended for the functional assays.29 The p.(Pro72His) variant, which affects the same amino acid as the common p.(Pro72Arg) PEX4 polymorphism (rs1042522), yielded a score of 6.1 with no detectable effect on p53 mRNA, suggesting that it has a moderate effect on p53 function. For this patient, an EBV-cell line was established, and the functional assay performed on the cell line yielded comparable results with a reduced score of 7.5 (normal score in EBV>10) and no impact on p53 mRNA levels. The p.(Pro191Arg) variant and the c.-117G>T variant had no detectable impact on the transcriptional activity, which led us to consider them as functionally normal.
Functional impact of the TP53 c.*1175A>C polymorphism located within the polyadenylation signal
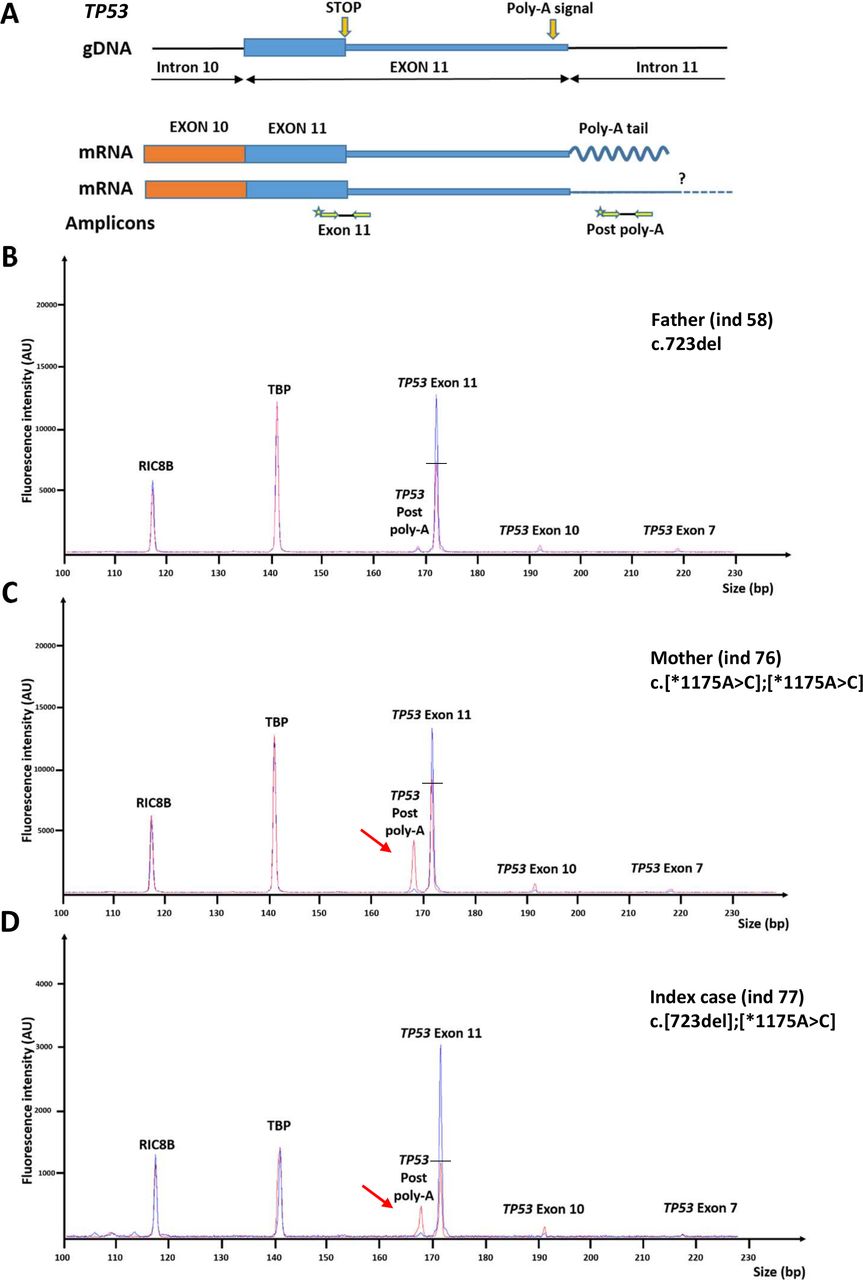
We performed the assay in the unaffected mother (individual 76, table 1 and online supplemental table 3) of a young female patient (individual 77, online supplemental table 3) who developed a high-grade glioma at 5 years of age and who carried a frameshift TP53 variant [c.723del, p.(Cys242Alafs*5)], transmitted by her father (individual 58, table 1 and online supplemental table 3). Unexpectedly, the mother had a reduced score of 5.5 and a moderate decrease in p53 mRNA levels (69%). A second blood sample yielded the same results (5.7; 73%). To explore this discrepancy, we reanalysed by NGS the whole TP53 locus in this family. This analysis revealed that the affected daughter carried the rs78378222 c.*1175A>C variant located within the polyadenylation signal, in addition to the frameshift TP53 variant. This polymorphic variant, present in 1.7% of non-Finnish Europeans, had previously been shown to be a risk factor for glioma and to impair 3′-end processing of p53 mRNA.30–33 Remarkably, the reanalysis of the TP53 locus in the mother revealed that she was homozygous for the c.*1175A>C variant. The p53 functional score in the index case harbouring the p.(Cys242Alafs*5) variant and the c.*1175A>C polymorphism (individual 77, online supplemental table 3) was lower than that obtained in her father harbouring only the frameshift variant (3.1 vs 7.1 and 6.0; see table 1 and online supplemental table 3). We analysed the impact of the c.*1175A>C variant on p53 mRNA 3’end processing using a dedicated RT-QMPSF with two amplicons located upstream and downstream of the polyadenylation site (‘exon 11’ and ‘postpoly-A’ respectively). This analysis revealed a reduction in p53 normal transcripts and the appearance of longer transcripts in the mother’s and daughter’s blood cells (figure 3).
{kind=link}
{kind=link}
{kind=link}
Impact of the heterozygous and homozygous TP53 c.*1175A>C variation on p53 pre-mRNA 3′ end processing. (A) Schematic representation of the TP53 3′ end region. The c.*1175A>C variant is predicted to yield at least two different transcripts; the upper one corresponds to the normal transcript with pre-mRNA cleavage and polyadenylation, and the lower one to longer transcript that extends after the poly-A signal. ‘Exon 11’ primers amplify both transcripts, while ‘postpoly-A’ primers specifically amplify the longer transcripts. As postpoly-A primers could also amplify gDNA, primers ‘exon 7’ and ‘exon 10’, which are specific to gDNA, were added to the reaction in order to monitor DNA contamination. (B) RT-QMPSF result obtained for the index case’s father (individual 58, S1; table 1 and online supplemental table 3) carrying the variant TP53 c.723del, p.(Cys242Alafs*5). The profile (in red) was superimposed on the profile of a control individual wild type for TP53 (in blue), using the control amplicons RIC8B and TBP. (C) RT-QMPSF result obtained for the index case’s mother (individual 76, S1; table 1 and online supplemental table 3) carrying the c.*1175A>C variant at the homozygous state. (D) RT-QMPSF result for the index case (individual 77, online supplemental table 3) carrying the c.723del, p.(Cys242Alafs*5) variant and the c.*1175A>C in trans. Red arrows indicate the appearance of longer p53 transcripts. The horizontal bars show the reduction of the normal p53 transcript level, as compared with the control. RT-QMPSF, reverse transcription–quantitative multiplex PCR of short fluorescent fragment.
Discussion
The interpretation of germline TP53 variants in patients with cancer is critical and should be performed before starting treatment considering their medical impact. The main objective of our assay was to provide a fast functional classification of rare uncharacterised variants in order to help clinicians with decision-making. Compared with the previous assay that we developed in EBV-immortalised lymphocytes,27 28 this blood assay does not require long-term cell culture and the results can be obtained within 1 week, fulfilling the timing required for diagnostic practice. The only constraint is to perform it within 48 hours after blood sampling in order to obtain robust results. Under these conditions, we were able to successfully analyse samples sent from other European countries.
Our assay fulfils most of the recommendations recently published by the Clinical Genome Resource Sequence Variant Interpretation working group regarding the clinical validity of functional assays29: (1) compared with the previously described p53 functional assays that test in vitro either cloned cDNA in yeast or artificial mutant libraries in cancer cell lines,23–25 this blood assay is performed in clinical samples in the patients’ genetic context; (2) the assay evaluates the transcriptional activity of p53 and not a specific domain of the protein; (3) it analyses simultaneously the impact of the variant on protein function and mRNA levels; (4) it was validated using 51 wild-type TP53 controls and 8 patients with seven distinct pathogenic or likely-pathogenic TP53 variants; and finally, (5) results show the robustness of the assay. Indeed, as shown in table 1, for 12 tested variants, we were able to perform the assay on EBV-immortalised cell lines and the results were very similar. Moreover, for five individuals, two different blood samples were tested and yielded similar results (table 1), and two variants (c.844C>T, p.(Arg282Trp); c.847C>T, p.(Arg283Cys)) were tested on two different individuals’ blood with concordant results (4.8 vs 5.0 and 5.3 vs 6.4).
We observed among the wild-type TP53 individuals a wide range of functionality scores (7.5–22.8). This probably suggests that there is a variability of the p53-mediated transcriptional response to DNA damage in the general population, although no obvious impact of age, clinical status or sex could be observed. The thresholds used in this study could be refined by testing additional deleterious variants. Despite this variability, all pathogenic/likely pathogenic variants generated low p53 functionality scores, and variants resulting in premature stop codons were also detected by a clear reduction of p53 mRNA levels. In addition, our assay allows testing of non-missense variants such as in frame indels. It should be highlighted that none of the previously published functional assays can be considered as a gold-standard method to classify germline TP53 variants.23–25 Therefore, no available p53 functional assay can be used to calibrate the blood assay. Indeed, as illustrated in table 1, discordant results were obtained for variants unambiguously classified in ClinVar as pathogenic or likely pathogenic. In particular, the founder Brazilian p.(Arg337His), an example of a variant with low penetrance, highlights the limits of the available tools. Whereas segregation data performed on large Brazilian pedigrees have clearly shown that this variant is pathogenic,34 bioinformatic predictions and functional analyses35 are conflicting (table 1). Our blood functional assay clearly shows that this variant alters the transcriptional activity of p53, although to a lesser extent than DNE missense variations, highlighting the limits of functional assays based on overexpression of cDNA. This result was confirmed in four additional patients carrying this variant using EBV cell lines (table 1).
The blood functional assay performed on PBMC harbouring unclassified variants led us to consider 12 variants (p.(Pro72His), p.(Gly105Asp), p.(Arg110His), p.(Phe134Leu), p.(Arg158Cys), p.(Pro278Arg), p.(Arg283Cys), p.(Leu348Ser), p.(Asp352Tyr), p.(Gly108_Phe109delinsVal), p.(Asn131del), p.(Leu265del)) as ‘functionally abnormal’, some with high impact. The interpretation is particularly challenging for p.(Pro72His), p.(Arg110His), p.(Arg158Cys), p.(Arg283Cys) and p.(Asp352Tyr) variants, as they were considered in yeast assays as functional or partially functional, and the Giacomelli assay classified them as not LOF_not DNE or was not conclusive. The low functionality score observed for p.(Arg110His) was confirmed in an EBV cell line derived from the patient and confirmed in two EBV cell lines from other patients carrying this variant. The result for the p.(Asp352Tyr) variant was confirmed on a second blood sample and with an EBV cell line derived from another patient also carrying this variant. The effect of p.(Arg283Cys) was also confirmed in EBV cell lines derived from the patient and from three additional patients with the same variant (table 1).
The clinical utility of the p53 functional assay is highlighted by the p.(Pro191Arg) variant. This variant was initially detected in a child with medulloblastoma at 2 years of age and whose brother died from a fibrosarcoma. Presymptomatic testing revealed that an unaffected brother (18 months), the mother and two maternal aunts were also carriers. We were then requested to evaluate this variant, and the functional assay performed in the maternal aunt (individual 65, online supplemental table 3) clearly showed that this variant does not alter the p53 transcriptional activity (table 1 and online supplemental table 3). Considering this result, segregation analysis was performed on the brother’s fibrosarcoma sample, revealing the absence of the variant and consolidating the conclusion of a non-pathogenic variant.
Our results show that this blood functional assay is also able to detect TP53 variations outside the coding regions, which are the only regions commonly analysed. Thanks to this assay, we discovered that the unaffected mother of an index case was homozygous for the polymorphic c.*1175A>C variant, and we show that this variant decreases p53 mRNA by altering the polyadenylation signal and produces longer transcripts extending beyond the poly-A site, as previously reported.30 When present on both alleles, this variant impacts p53 functionality with the same magnitude as a germline pathogenic TP53 variant. This prompted us to recommend breast MRI every year for this unaffected adult relative. We had the opportunity to perform the assay on EBV-immortalised lymphocytes harbouring only this heterozygous variant, and we observed a normal score (data not shown), suggesting that the heterozygous c.*1175A>C variant alone is insufficient to alter p53 function. The comparison of the p53 functional scores observed in the index case who developed a high-grade glioma at 5 years of age and harbours the null c.723del, p.(Cys242Alafs*5) variant and in trans the polymorphic c.*1175A>C variant, and in her father carrying only the TP53 null variant suggests that the c.*1175A>C variant may act as a genetic modifier in pathogenic TP53 variant carriers and could increase the risk of glioma in carriers, as previously shown in the general population.30–33
In summary, we suggest that our blood p53 functional assay should be a useful tool not only for the rapid interpretation of germline TP53 variants of unknown significance in clinical practice, in complement to the previously developed assays, but also for the indirect detection of cryptic alterations within regulatory regions impacting p53 function.
Data availability statement
All data relevant to the study are included in the article or uploaded as supplementary information. Deidentified participant data are available from thierry.frebourg@chu-rouen.fr.
Ethics statements
Patient consent for publication
Acknowledgments
The authors are grateful to their French and European colleagues for providing clinical information and sending blood samples for TP53 analysis. The authors are indebted to Philippe Ruminy (Inserm U1245, Comprehensive Cancer Centre Becquerel, Rouen) for advices on the reverse transcription–multiplex ligation probe amplification experiments and to Nikki Sabourin-Gibbs (Rouen University Hospital) for her assistance in editing the manuscript.
References
Supplementary materials
Supplementary Data
This web only file has been produced by the BMJ Publishing Group from an electronic file supplied by the author(s) and has not been edited for content.
Footnotes
SR and MR contributed equally.
Contributors IT, GB and TF formulated research goals and aims. Functional analyses were performed by SR and MR. RNA-Seq was performed by SR, CD and IT and bioinformatic analysis by RL and SC. Molecular analyses were performed by JB, EB, FC, GL, SV and supervised by GB, EK and SBD. TF, IT and GB supervised the overall progress and manuscript writing. SR and MR contributed equally to this work.
Funding This work was supported by the North-West Canceropole, the European Regional Development Fund (European Union and Région Normandie), the Association pour la Recherche contre le Cancer and the Ligue Contre le Cancer.
Competing interests None declared.
Provenance and peer review Not commissioned; externally peer reviewed.