Article Text

Statistics from Altmetric.com
- BPES, blepharophimosis-ptosis-epicanthus inversus syndrome
- CAI, codon adaptation index
- Dpc, days post-coitum
- ORF, open reading frame
- PAGE, polyacrylamide gel electrophoresis
- POF, premature ovarian failure
Mutations in the FOXL2 gene have recently been shown to cause the blepharophimosis-ptosis-epicanthus inversus syndrome (BPES), a rare genetic disease (MIM 110100).1 In type I BPES eyelid abnormalities are associated with premature ovarian failure (POF), while in type II BPES only the eyelid malformation is observed.2FOXL2 is the first human autosomal gene whose dominant mutations have been shown to be involved in POF. The eyelid malformation in both BPES types is inherited as a dominant trait and we have recently argued that ovarian failure in type II BPES is a recessive trait.3
FOXL2 is a single exon gene of 2.7 kb. The predicted protein of 376 amino acids belongs to the large family of forkhead/winged helix transcription factors, containing a characteristic 100 amino acid DNA binding forkhead domain. Many members are known to be involved in vertebrate embryogenesis4 and some have been implicated in inherited developmental human disorders.5 Apart from the forkhead domain, the FOXL2 protein also contains a polyalanine (poly-Ala) tract whose role has not yet been elucidated. Recurrent mutations leading to its expansion cause type II BPES and account for 30% of the deleterious alterations detected in the open reading frame (ORF).6,7 These alleles have been considered as hypomorphic (residual activity) in the context of the ovary.1,6
Northern blot analysis and RNA in situ hybridisation have shown that FOXL2 is expressed in developing mouse eyelids and in adult ovarian follicular cells.1
Here we have performed a comparative sequence analysis of FOXL2 in order to study the evolution of the FOXL2 coding region. We have estimated the synonymous (Ks) and non-synonymous (Ka) substitution rates in the ORF of several species, human, goat, mouse and pufferfish. In addition, to determine the subcellular localisation of the FOXL2 protein and its expression pattern, we have developed and characterised two anti-peptide polyclonal antibodies.
In summary, we showed that FOXL2 orthologues are highly conserved and code for a nuclear protein specifically expressed in fetal eyelids and in both fetal and adult ovarian follicular and stromal cells. Ovarian FOXL2 expression begins at early stages of development, before the onset of folliculogenesis. FOXL2 is the earliest known sex dimorphic marker of ovarian differentiation in mammals and may play a multiple role during fetal development and also later, during adult female fertile life.
MATERIALS AND METHODS
Comparative sequence analysis
To obtain the full length mouse sequence, we directly sequenced by primer walking8 the BAC 13L19 (RP23 Mouse BAC Library, Research Genetics), which contains the whole mouse foxl2 locus (GenBank AI: AF522275). Human, rat, and Fugu FOXL2 sequences were assembled from GenBank (AF301906, AC105826.1, Scaffold_8165/ProtJGI_24134). Goat FOXL2 sequence was obtained from two genomic clones (Genbank AI: AY112725).
Comparative sequence analysis was performed using the method of Li9 implemented in the DAMBE package10; for each pairwise comparison, the 0-fold, 2-fold, and 4-fold degenerate sites were counted. The two sequences were then compared codon by codon, and the number of transitions and transversions at each of the three categories of sites were counted. The two parameter method of Kimura11 was then applied to correct the number of transitions and transversions for multiple hits. Then, synonymous and non-synonymous substitutions were calculated as described previously.9
Key points
-
Mutations in FOXL2, a forkhead transcription factor gene, have recently been shown to cause the blepharophimosis-ptosis-epicanthus inversus syndrome (BPES). This rare genetic disorder leads to a complex eyelid malformation associated or not with premature ovarian failure (BPES type I or II, respectively).
-
We performed a comparative analysis of the FOXL2 sequence in several species (human, goat, mouse, and pufferfish) showing that the FOXL2 coding region is highly conserved in these species.
-
The FOXL2 protein contains a polyalanine tract whose role has not yet been elucidated. Recurrent mutations leading to its expansion result in BPES type II and account for 30% of the deleterious alterations detected in the open reading frame (ORF) of FOXL2. We showed that the number of alanine residues is strictly conserved among the mammals studied, suggesting the existence of strong functional or structural constraints.
-
We provide immunohistochemical evidence indicating that FOXL2 is a nuclear protein specifically expressed in eyelids and in fetal and adult ovarian follicular cells. It does not undergo any major post-translational maturation.
-
FOXL2 is the earliest known marker of ovarian differentiation in mammals and may play a role in ovarian somatic cell differentiation and in further follicle development and/or maintenance.
Immunochemistry
Two custom made polyclonal antibodies directed against the most N- and C-terminal hexadecapeptides corresponding to the human FOXL2 protein were produced in the rabbit (Eurogentec. Peptides: N-MMASYPEPEDAAGALL, C-WDHDSKTGALHSRLDL).
Protein extraction
Goat ovaries were homogenised in RIPA buffer (150 mmol/l NaCl, 10 mmol/l Tris-HCl, pH 7.4, 1% Nonidet P40, 5 mmol/l EDTA, 0.1% SDS) containing a cocktail of protease inhibitors. Human extracts were obtained from Clontech (ovary protein medley). Twenty-five μg of each protein extract denatured in Laemmli sample buffer were electrophoresed and electrotransferred onto nitrocellulose membranes (Novex). Immunoblots were performed following standard procedures (first antibodies diluted 1/1000; 1/500 for preimmune serum; incubation overnight at 4°C; secondary antibody, anti-rabbit IgG-peroxidase (Perbio/Pierce) diluted 1/10000; incubation 45 minutes at room temperature). Enhanced chemiluminescence detection was carried out as described by the manufacturer (Amersham).
Immunoprecipitations
A goat ovary was homogenised in RIPA buffer (300 mmol/l NaCl) and inhibitors without detergent (protein concentration 15 μg/μl). Then immunoprecipitation was performed following standard procedures (pre-clearing: one hour with 1/100 preimmune serum, then one hour with protein A-sepharose (Amersham); precipitation: anti-N or anti-C antibodies diluted 1/100, 4°C, overnight). Extracts precipitated with anti-N antibody were detected with the anti-C antibody and vice versa.
Immunohistochemistry
For human tissues, paraffin embedded sections were treated by standard means and the anti-FOXL2 antibodies diluted 1/1000. For goat and mouse ovaries, 7 μm cryosections were fixed in 4% paraformaldehyde, and the anti-C antibody was diluted 1/1500. In both cases, primary antibodies were detected by indirect immunohistology using peroxidase/diaminobenzydine (Perbio/Pierce). The Necker Hospital Anatomopathological Service (Paris) follows the ethical standards of the centre
FOXL2 expression in bacteria
The FOXL2 ORF to be expressed in E coli was amplified from human genomic DNA by PCR (Vent polymerase, 10% dimethylsulphoxide) and primers pBAD-FOXL2F: 5′-ATGATGGCCAGCTACCCCGAG-3′ and pBAD-FOXL2R: 5′-GAGATCGAGGCG CGAATGCAGC-3′. After adenylation of the amplicon (Taq polymerase) it was cloned using the Invitrogen pBAD expression system. The accuracy of the ORF sequence was checked by sequencing. Induction of FOXL2 expression was carried out following the manufacturer’s instructions (Invitrogen).
RESULTS AND DISCUSSION
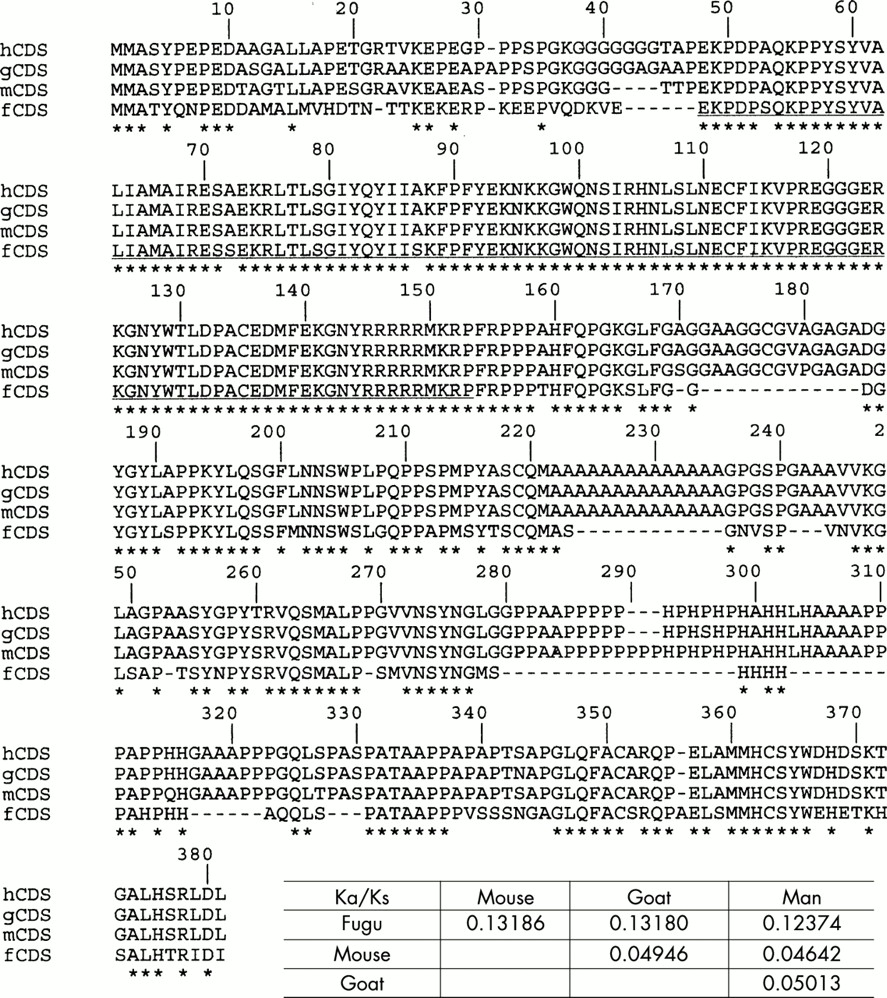
To study the pattern of evolution of FOXL2 and to find out the type of selection operating on the coding region, we compared its sequence in human, goat, mouse, and Fugu. We used the method of Li,9 which consists of estimating the number of synonymous substitutions per synonymous site (Ks) and the number of non-synonymous substitutions per non-synonymous site (Ka), as the ratio Ka/Ks is a good indicator of the selective pressure at the protein level.12 Ka/Ks <1 is suggestive of strong (dominantly purifying) selection whereas a Ka/Ks >1 indicates a diversifying positive selection.9,12 Amino acid sequences were found to be very similar among mammals (above 96%), which was confirmed by very low Ka/Ks ratios obtained for the whole ORF, suggesting that FOXL2 is under purifying selection. A similar comparative analysis carried out on Fugu FOXL2 showed that the forkhead domain and the C-terminal region were highly similar to those of the other species studied, whereas the N-terminal region was more divergent and the poly-Ala tract was absent (fig 1). The forkhead and C-terminal regions may have conserved their functions through evolution, while the N-terminal region would evolve under weaker constraints. The difference between Ks and Ka is less dramatic in comparisons between the Fugu sequence and the three mammalian sequences. This may simply reflect that Ks is underestimated between highly diverged sequences (multiple hits that erase each other). We have also studied FOXL2 properties in silico. The calculated isoelectric points of all orthologues are above 9.7, consistent with the putative DNA binding activity of FOXL2. Hydropathy plots of all orthologues are also very similar (data not shown). These results confirm the conservation of FOXL2 and its properties during evolution.

Comparative sequence analysis of FOXL2 orthologues (performed as in Li9 using the DAMBE package10). The Ka/Ks ratios are reported in the table. Asterisks represent fully conserved sites. Amino acid sequences are very similar among mammals (above 96%) as confirmed by very low Ka/Ks ratios obtained for the whole ORF. The forkhead domain (underlined) and the C-terminal region (excluding gaps) of Fugu are very conserved while the N-terminus is more divergent. Notice that the polyalanine tract is absent in Fugu but its length is extremely conserved among mammals (including rat, not shown). It is clear that in mammals the length of the tract is under strong constraints. Other gaps present in the Fugu sequence might reflect the general tendency of its genome to compaction.
The poly-Ala tract present in the mammalian orthologues is very likely the result of a GCN expansion (fig 1) and as pointed out above its unscheduled extension leads to hypomorphic deleterious alleles in man that account for 30% of intragenic FOXL2 mutations.6,7 Poly-Ala tracts generally form alpha-helices with multiple isoforms and their expansion may lead to protein misfolding.13 To date, six other human diseases have been associated with a deleterious expansion of a poly-Ala tract (reviewed in OMIM 300382). Since FOXL2 poly-Ala expanded proteins behave as hypomorphic alleles in the context of the ovary (BPES type II), it is very likely that a proportion of the abnormal protein is still functional. However, one cannot rule out that the inactive fraction may aggregate forming intranuclear inclusions (as reported in oculopharyngeal muscular dystrophy, MIM 164300).14 The number of alanine residues (14) is strictly conserved among the mammals studied (including rat, data not shown). This suggests the existence of a strong functional or structural constraint so that an expansion of the tract results in a deleterious effect.
To determine the subcellular localisation and the expression pattern of FOXL2, we have produced two polyclonal antibodies directed against FOXL2, able to recognise the mammalian orthologues. Western blot analyses carried out with each antibody on human and goat ovary protein extracts showed a 45 kDa specific band corresponding to the FOXL2 protein (fig 2). We also performed cross-immunoprecipitations using ovary protein extracts (immunoprecipitation with one anti-FOXL2 antibody and detection with the other one). A 45 kDa band was consistently obtained in both cases (fig 2). The FOXL2 signal on western blots was strong. This is compatible with in silico estimations of the codon adaptation indexes (0<CAI<1) of the ORFs, a measure that correlates positively with the level of expression. For all ORFs the CAI were above 0.7, which is suggestive of high expression levels. Finally, FOXL2 was expressed in E coli. Using either the anti-N or the anti-C antibody on extracts from induced and non-induced bacteria, a specific 50 kDa band was detected in the induced extracts (fig 2), which corresponds to the FOXL2 protein (and c-myc and polyhistidine tags). This further showed the specificity of the antibodies. Besides, the fact that we obtained a band of similar size as that detected in mammals suggests that FOXL2 does not undergo any gross post-translational maturation.
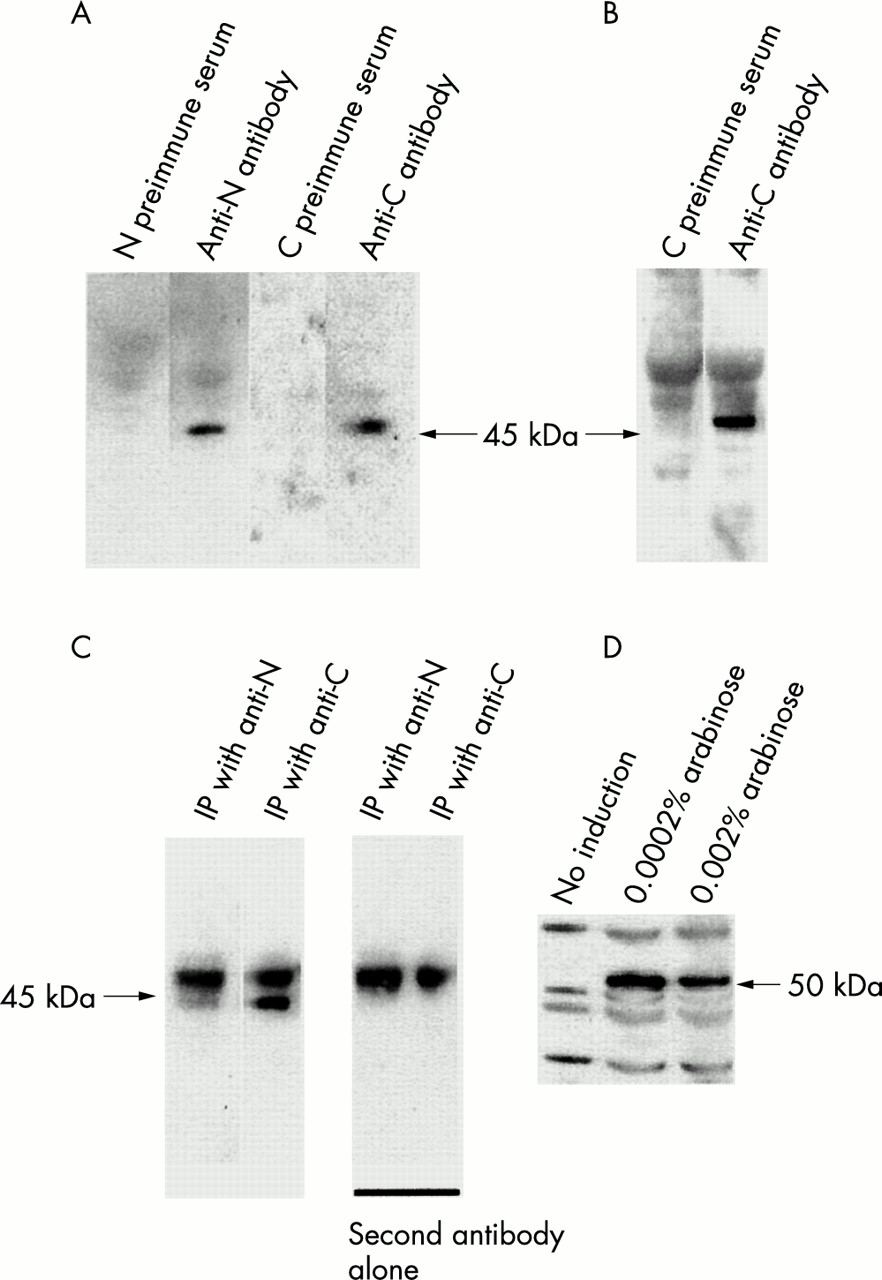
Antibody characterisation. Protein extracts were blotted and detected with pre-immune sera or anti-sera as indicated (4-12 % bis-tris SDS-PAGE, Novex). (A) Goat ovary protein extract. (B) Human ovary protein medley. (C) Immunoprecipitations (IP) followed by immunoblots. First two lanes: immunoprecipitation with anti-N or anti-C antibodies and detection with anti-C or anti-N antibodies respectively. Remaining lanes: negative controls with secondary antibody alone (staining the IgG heavy chains). (D) Expression of FOXL2 in E coli detected with the anti-C antibody without or with arabinose induction (Invitrogen pBAD system). The 50 kDa specific band corresponds to FOXL2 plus the c-myc and polyhistidine tags. This suggests that FOXL2 does not undergo any major post-translational modification.
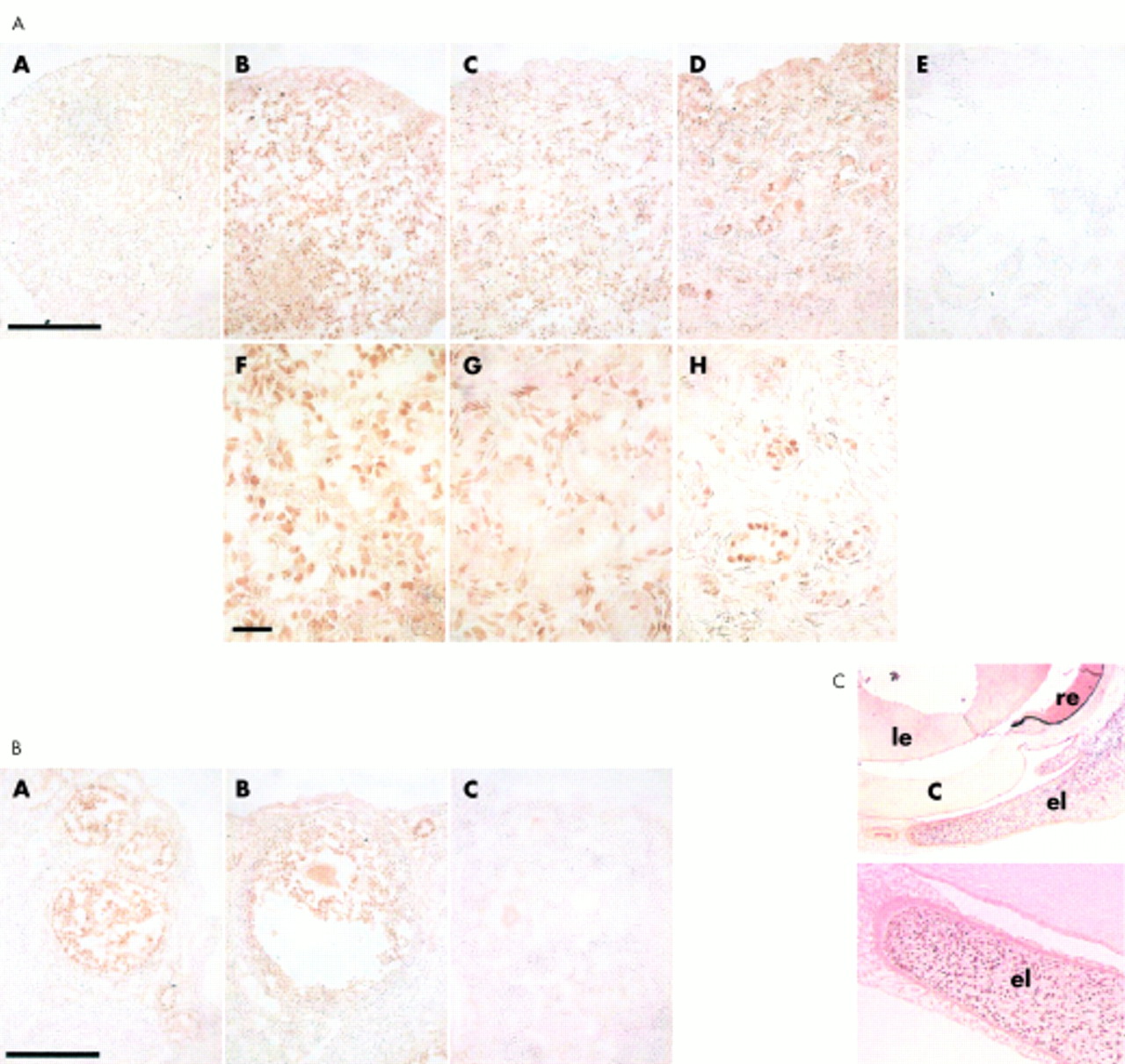
Immunohistochemical experiments showed that FOXL2 was a nuclear protein specifically present in fetal and adult ovaries and eyelids (figs 3 and 4). All other human and goat fetal tissues studied (heart, pancreas, testis, digestive tract, thymus, thyroid, kidney, spleen, uterus) including goat embryo sections were devoid of specific FOXL2 signal. The specific nuclear localisation of FOXL2 is consistent with its putative role as a transcription factor. In the human, FOXL2 was shown to be expressed in a population of cells of the female genital ridge before sex determination (fig 3). In fetal ovaries (10 weeks of fetal life), a similar pattern was observed. At these stages, no clear structural organisation of the gonad is detectable by histology. At later stages (22 and 29 weeks) FOXL2 was highly expressed in follicular cells but also in stromal cells. At 40 weeks of fetal life and in the adult ovary this pattern was maintained (fig 3). This contrasts with previously reported in situ hybridisation data in mouse showing ovarian mRNA expression exclusively in follicular cells.1

Immunohistochemistry performed with anti-C antibody on human ovary sections from different stages. (A) Female genital ridge (before sex determination). (B) Fetal ovary at 10 weeks, (C, D) at 22 weeks, (E, F) at 29 weeks, (G, H) at 40 weeks (final developmental stage). (I, J) 16 year old ovary. (K) Graafian follicle. Scale bars correspond to 10 μm in B, D, F, H, and J, to 200 μm in C, E, and I, and to 150 μm in G. FOXL2 expression begins at an early stage of development (genital ridge). At 22 weeks of development, follicles are visible. These round structures (indicated by arrows) are composed of oocytes surrounded by follicular cells (granulosa cells). FOXL2 is highly expressed in follicular cell nuclei and also (though more weakly) in stromal cell nuclei. Fig 3K shows a Graafian follicle which contains two types of follicular cells, mural granulosa cells (outer densely stained layer, labelled “mg” on the figure) and cumulus granulosa cells (directly surrounding the oocyte, “cg”). Both types express FOXL2. Note, in the low magnification images (C, G, E, and I), how the density of follicles tends to decline with developmental time. FOXL2 mutations are thought to accelerate this process.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
FOXL2 protein expression during goat eyelid and gonadal development and in mouse adult ovary. (Section A) FOXL2 protein was not detected in goat ovaries at 32 dpc (A) but was present in somatic cells of the female gonads at 36 dpc (B, F). Positive cells were located in all the gonad except the somatic cells of the surface epithelium and germ cells. At 56 dpc (C, G) the stained cells delineated the ovigerous cords that contain the germ cells. At 90 dpc (D, H) FOXL2 is highly expressed in follicular cells nuclei. Testis at 90 dpc was negative (E). In mouse adult ovary (section B), FOXL2 was only detected in follicular cells (A, B) but not in control without the primary antibody (C). This confirms the results obtained by Crisponi et al1 using in situ hybridisation.1 Scale bars correspond to 100 μm in section A (A, B, C, D, E) and section B (A, B, C) and to 10 μm in section A (F, G and H). (Section C) Immunostaining of a 45 dpc fetal goat eye section with anti-C antibody. FOXL2 is strongly expressed in the developing eyelids whereas others tissues are negative. el, eyelid; c, cornea; le, lens; re, retina.
In order to study the developmental expression of FOXL2 better, we performed similar experiments on fetal caprine gonads (explored stages: 32 days post coitum (dpc), 36 dpc, 56 dpc, 90 dpc). The goat is a well known model because of the existence of an XX sex reversing mutation that maps near the FOXL2 locus.15 Similar to human and in contrast to mouse, this species is mono-ovulating. The earliest stage where the FOXL2 protein was detected (36 dpc) corresponds to the time of Sertoli cell differentiation in testis16 (fig 4). At this stage, no clear structural organisation was visible in the ovaries. Whatever the stage of development, the FOXL2 protein was specifically detected in ovaries while testes were negative. At 56 dpc, when female meiosis begins, the stained cells delineated the ovigerous cords containing the germ cells. Then, when the formation of primordial follicles occurs (90 dpc), the most positive cells corresponded to follicular cells.
Previous studies using quantitative RT-PCR and northern blotting have shown that FOXL2 mRNA is present in the goat at 30 dpc in the mesonephros and/or gonads.15 Besides, FOXL2 mRNA expression was detected in testes at several developmental stages and even increased beyond 70 dpc. Here, no FOXL2 protein could be detected in any of those tissues, at the developmental times mentioned, suggesting that FOXL2 undergoes post-transcriptional or translational regulation.
In conclusion, our data show that FOXL2 expression is specific to eyelids and ovaries in mammals, which is compatible with the BPES phenotype. FOXL2 is the earliest known marker of ovarian differentiation in mammals, may undergo post-transcriptional or translational control, and may play a role at an early stage of development in ovarian somatic cell differentiation. As it is still strongly expressed in postnatal and adult follicular cells, it may also play a role during all female fertile life in follicle development and/or maintenance.
Acknowledgments
We thank J F Alkombre for animal care and B Vigier for the histology.
REFERENCES
Footnotes
-
↵* Present address: U361 Reproduction et Physiopathologie Obstétricale, Hôpital Cochin, Pavillon Baudelocque, 123 Bd de Port-Royal, 75014 Paris, France