Article Text

Statistics from Altmetric.com
- HDGC, hereditary diffuse type gastric cancer
- HNPCC, hereditary non-polyposis colorectal cancer
- CHO, Chinese hamster ovary
- DHPLC, denaturing high pressure liquid chromatography
Germline mutations of the cell adhesion molecule E-cadherin have been shown for the first time to underlie a hereditary diffuse type gastric cancer syndrome (HDGC) in Maori families,1 and subsequently have been reported in HDGC patients of various ethnic origins.2–9 Gastric cancer may also be associated with other hereditary tumour syndromes, which are mainly characterised by carcinomas of other organs. One of these syndromes is the HNPCC syndrome (hereditary nonpolyposis colorectal cancer syndrome), which is caused by germline mutations in DNA mismatch repair genes.10 Furthermore, gastric cancer has been observed in the context of the Li Fraumeni syndrome, a rare cancer syndrome due to germline mutations of the TP53 tumour suppressor gene,11–14 as well as in association with the FAP and Peutz–Jeghers syndrome.15
Despite known molecular genetic causes, contributing to a genetic predisposition to gastric cancer, a considerable number of familial cases have been reported that were not attributable to one of this hereditary syndromes, suggesting that unknown susceptibility genes for gastric cancer might exist. Putative tumour suppressor genes that are commonly inactivated in sporadic gastric cancers, such as the RUNX3 or HPP1 genes, are potential candidate susceptibility genes.16,17
In a previous study, we identified one E-cadherin germline mutation among seven diffuse type familial gastric cancer patients, indicating that in addition to E-cadherin, other genes might be involved in genetic predisposition to the disease in this patient group.4
There were three goals of the present study. First, we aimed to extend our analysis of E-cadherin germline mutations to a higher number of patients, to evaluate the contribution of germline mutations in this gene to German familial and early onset gastric cancer patients and to characterise identified missense mutations for their functional relevance. Secondly, we wished to analyse the role of TP53 germline mutations for a familial aggregation of the disease and for early onset patients. Our third purpose was to investigate two putative tumour suppressor genes, RUNX3 and HPP1, for their potential role in genetic predisposition to gastric cancer.
MATERIALS AND METHODS
Definition of familial gastric cancer
Criteria for the definition of familial gastric cancer are: at least two first or second degree relatives with gastric cancer, one diagnosed before the age of 50; or at least three first or second degree relatives with gastric cancer, independent of age at diagnosis. Gastric cancer of the index patients was confirmed by histopathological reports.
The study protocol was reviewed and approved by the local ethics committee, and DNA samples and family history were obtained with the informed consent of the patients.
Key points
-
The aim of the study was to investigate the role of E-cadherin germline mutations in a series of German familial and early onset gastric cancer patients, and to evaluate three other genes, TP53, RUNX3, and HPP1, which are involved in gastric carcinogenesis, for their potential role in a genetic predisposition to the disease.
-
Two E-cadherin germline mutations, one truncating and one novel missense mutation, were found among 30 (7%) familial gastric cancer patients with diffuse or mixed type gastric cancer. Functional analysis of the missense mutation demonstrated alterations of cell adhesion and cell invasion, which strongly support its pathogenic role. One novel truncating mutation was identified among the 15 (7%) early onset gastric cancer patients. The mother of this patient was affected with bilateral breast carcinoma, which underlines the association of breast carcinoma with germline mutations in the E-cadherin gene.
-
Mutation analysis of the TP53 gene revealed one germline mutation among 34 (3%) familial gastric cancer patients, indicating that the TP53 gene should be included in the genetic screening of familial gastric cancer patients with a suspected genetic predisposition to the disease.
-
No mutations were identified in the RUNX3 and HPP1 genes, suggesting that these are not major gastric cancer predisposition genes.
Patients for germline mutation analysis
DNA samples from 35 familial gastric cancer patients from Germany were analysed for germline E-cadherin mutations; seven of these patients were included in a previous study.4 Of the index patients, 24 were affected with diffuse type, six with mixed type, and five with an intestinal type of gastric cancer according to the classification of Lauren.18 Four of the 24 families fulfilled the strict criteria for HDGC, defined by the International Gastric Cancer Linkage Consortium (IGCLC).19 In these four families, besides the histological verification of the index case, the diagnosis was confirmed in at least another family member (table 1).
Summary of E-cadherin germline mutation analysis
In addition to the 35 families showing aggregation of gastric cancer, 15 early onset patients, with gastric cancer diagnosed before the age of 45 years, were included in this study. Of these patients, 14 had a diffuse type and one a mixed type gastric carcinoma (table 1).
Germline mutation analysis of the TP53, RUNX3, and HPP1 genes was performed for 34 German familial gastric cancer patients, 20 with diffuse type, five with mixed type, and nine with intestinal type of gastric cancer, according to the classification of Lauren (table 2).18 For mutation analysis of the HPP1 gene, one Portuguese and one Canadian family with gastric polyposis and diffuse type gastric cancer20 were included in this part of the study. The 15 early onset gastric cancer patients were analysed for germline mutations in the TP53 gene, but not in the RUNX3 and HPP1 genes. Gastric cancer patients related to HNPCC due to germline mutations in the DNA mismatch repair genes hMLH1 or hMSH2 were not included in this study.
Results on TP53 germline mutation analysis in familial gastric cancer
Mutation analysis
Genomic DNA was isolated from peripheral lymphocytes by phenol chloroform extraction or by the flexigene DNA extraction kit (Qiagen, www1.qiagen.com) according to the manufacturer’s instructions. DNA from paraffin embedded tumour tissue was isolated after manual microdissection as described previously.4
Mutation analysis was performed by DHPLC (Wave System, Transgenomic, Omaha, NE, USA) of the PCR products of all coding exons, including the intron-exon boundaries of the corresponding genes. Primers for the amplification of all 16 coding exons of the E-cadherin gene were according to published primer sequences.21 For TP53, exons 2–11 were analysed using primers, PCR, and DHPLC conditions as previously described.22,23 The five coding exons of the RUNX3 gene were amplified according to published primer sequences,16 with the exceptions of exon 1 and exon 5, for which modified primer sequences were used. Primer sequences for the 10 coding exons of the HPP1 gene were based on the DNA sequence of a chromosome 2 genomic contig containing the HPP1 gene (NT005403, gi29794150, NCBI data bank). The primer sequences and detailed PCR conditions are available from the authors on request.
The PCR reactions were performed in 50 μl of reaction mixture consisting of 10 mM Tris-HCl, pH 8.3; 50 mM KCl; 1.0, or 1.5, or 2.0 mM MgCl2; 0.01% gelatine; 200 µM each dNTP; 0.4 µM of each primer; and 2.5 U Taq Polymerase.
The optimal temperatures for resolution of heteroduplex and homoduplex DNA for the exons of the E-cadherin, RUNX3, and HPP1 genes were established by analysing the melting behaviour of the PCR fragments in the temperature range corresponding to the calculation of the Wavemaker™ software. The analysis temperature for each fragment was the point at which at least 75% of the DNA was present as an alpha helix, or 1–2°C higher. Fragments with more than one melting domain were analysed at additional temperatures. DHPLC conditions for each exons are available from the authors on request.
DNA sequence analysis
PCR products with an aberrant DHPLC chromatogram were directly sequenced in both directions, starting from a new PCR product, as previously reported.4 Samples demonstrating a putative pathogenic mutation were confirmed by another independent PCR and sequencing reaction.
Immunohistochemistry
p53 immunohistochemistry of the tumour tissue of the patient with the TP53 germline mutation was performed using the DO7 monoclonal antibody (Dako, www.dakocytomation.com) as described previously.22 E-cadherin immunohistochemistry of selected cases was performed with a monoclonal E-cadherin antibody (Clone 36, Transduction Laboratories, Hamburg, Germany) as described.5 Appropriate positive and negative controls were included with each staining series.
Functional assays
As previously described,24 construction of the plasmids encoding the E-cadherin mutants corresponding to the DNA sequence variants G1774A and C2396G were obtained by nested PCR, using wild type E-cadherin cloned into pcDNA3 vector as DNA template and primers carrying the desired mutations (G1774A: For 5′-gac aac acc ccc ata cca g -3′; Rev: 5′- at ggg ggt gtt gtc att cac -3′; For: ctc atg agt gtc ccc cgg tat ctt cgc -3′; Rev: 5′- gcg aag ata ccg ggg gac act cat gag -3′). Plasmids were stably transfected into Chinese hamster ovary (CHO); DG44 dhfr− cells and single emerging clones expressing equal amounts of E-cadherin protein were selected for the functional characterisation. According to a previous report,24 cell-cell adhesion was evaluated in a slow aggregation assay. Briefly, 2×10+4 cells were seeded on top of a matrix of soft agar (Bacto-agar, Difko Laboratories), evaluating aggregates formation at 24 hours and 48 hours with a phase contrast microscope. For the invasion studies, single cell suspensions were seeded on top of Matrigel Invasion Chambers (BD Biosciences, www.bdbiosciences.com) and the ability of cells to invade was evaluated according to the manufacturer’s instructions. In all experiments, at least two independent clones were used in order to exclude clonal dependence of the observed result.
RESULTS
E-cadherin mutation analysis
In the 35 German familial gastric cancer patients analysed so far, we found two germline mutations in the E-cadherin gene. Results in comparison with the histopathological type of the index patient are summarised in table 1. One mutation was a novel missense variant represented by a heterozygous C>G change at nucleotide position 2396, leading to an amino acid exchange from prolin to arginin at codon 799 (P799R). This was not found in an analysis of 50 control individuals, but was found in a 41 year old patient with a mixed adenocarcinoma of the oesophageal-gastric junction. Immunohistochemical analysis of E-cadherin expression in the tumour of this patient revealed a positive, membranous staining pattern in the tumour cells. DNA sequencing of exon 15 in the tumour revealed that the mutation was present in a heterozygous state, indicating that there was no LOH. There were two affected second degree relatives with gastric cancer in the maternal line (fig 1A); no samples were available from these family members for mutation testing.

Pedigree of the families with novel E-cadherin germline mutations. (A) Pedigree of the familial gastric cancer patient with the P377R mutation. (B) Pedigree of the early onset gastric cancer patient with the insertion G at nucleotide 1619. General symbols: squares, males; circles, females; numbers, age in years at diagnosis; arrow, index patient.
The other mutation found in the familial group was a truncating mutation in exon 3, described in our previous study.4
One novel truncating mutation was detected among the 15 early onset patients (14 with diffuse type and one with mixed type gastric cancer). The mutation was an insertion G at nucleotide 1619 in exon 11, leading to a frameshift with a premature stop at codon 547 (table 1). The patient had diffuse type gastric cancer at the age of 29. No family history of gastric cancer was elicited, but the mother had bilateral breast carcinoma at the age of 49, an abdominal tumour was reported for the maternal grandmother, and lung cancer for the maternal grandfather (fig 1B). Immunhistochemical analysis of E-cadherin expression in this case showed positive but diffuse cytoplasmatic staining in the tumour, where this mutation was also detected in an heterozygous state. However, we found an additional somatic E-cadherin mutation, an A>T transversion at nucleotide 1373 leading to a change from asparagin to isoleucin at codon 458 (D458I).
An unclassified sequence variant in intron 4 (532–18 C>T) was found in two familial diffuse type gastric cancer patients and in one of the 50 control individuals. A missense variant A592T (1774G>A) in exon 12 was identified in one familial gastric cancer patient and in one control. This missense variant was found in the patient who also carried a TP53 germline mutation and, in addition, several other known polymorphisms were found.
To address the pathogenic role of the two E-cadherin missense variants A592T and P799R, soft-agar aggregation and matrigel invasion assays were performed on CHO stably transfected cells, in comparison with cells expressing the wild type protein. Cells expressing the E-cadherin variant A592T were not invasive into Matrigel and gave rise to aggregates when seeded on top of an agar matrix, resembling the behaviour of cells expressing the wild type E-cadherin. These results reinforce the hypothesis that A592T is non-pathogenic. Cells expressing the E-cadherin missense variant P799R failed to aggregate and invade into Matrigel, suggesting that P799R pathogenic effect. Results are summarised in table 3.
Summary of the aggregation and invasion assays
TP53 mutation analysis
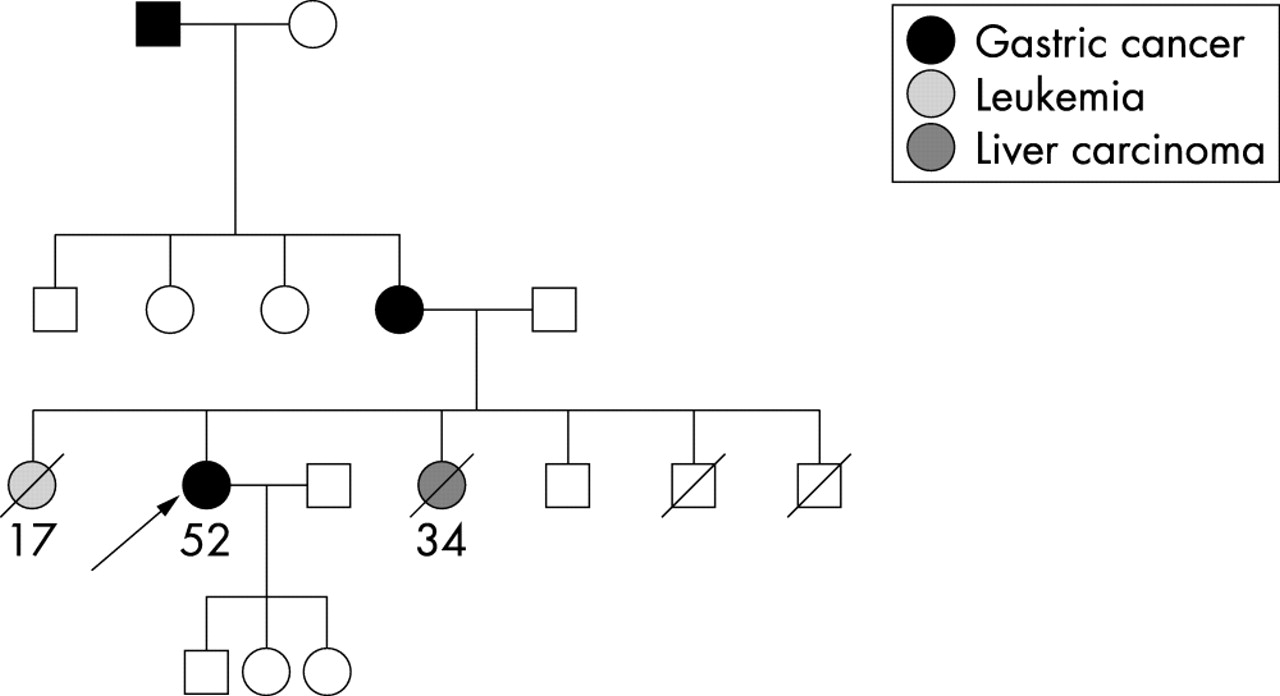
In one of 34 (3%) familial gastric cancer patients, a germline mutation in the TP53 gene was detected (table 2). The mutation was a heterozygous C>T transition at nucleotide 847 in exon 8, leading to a change from arginine to cysteine at codon 283 (R283C). This patient had a diffuse type gastric carcinoma, diagnosed at the age of 52. In the family there were three affected family members with gastric cancer. In addition, one sister of the patient died wwith leukemia at the age of 17 years, and another wth liver carcinoma at the age of 34 years (fig 2). No samples from these family members were available to test for the TP53 mutation. Mutation analysis of exon 5–8 of the tumour DNA showed that the mutation was present in a heterozygous state, but a somatic missense mutation in exon 5, a T>A transversion at nucleotide 389 resulting in an amino acid exchange from leucin to histidin at codon 130 (L130H), was found. Immunohistochemical analysis demonstrated positive TP53 expression in the tumour cells, but not in the adjacent non-tumorous epithelial cells.
{kind=link}
{kind=link}
Pedigree of the familial gastric cancer patient with the TP53 germline mutation R283C. General symbols: squares, males; circles; females; numbers, age in years at diagnosis; arrow, index patient.
No TP53 germline mutations were identified among the 15 early onset gastric cancer patients. A T>A transversion at nucleotide 1100+30 was identified in intron 10 in one familial and in one early onset gastric cancer patient, and was also found in one of the 50 control subjects. Several common polymorphisms, which have been already described, were also found.
RUNX3 mutation analysis
No germline mutations or polymorphisms were found in the RUNX3 gene in an analysis of 34 familial gastric cancer patients.
HPP1 mutation analysis
No germline mutations were detected in the HPP1 gene among the 34 familial gastric cancer cases from Germany, nor in one of the two gastric cancer families with a gastric polyposis phenotype, from Portugal and Canada. Two intronic DNA sequence variants, in intron 4 at nucleotide 439+26G>C and at nucleotide 439+47T>A, were detected in one patient and in two patients, respectively.
DISCUSSION
In this study we describe a comprehensive mutation analysis to elucidate the role of four different genes, E-cadherin, TP53, RUNX3, and HPP1, in genetic predisposition to gastric cancer. With respect to E-cadherin, we identified three germline mutations among the 35 familial and the 15 early onset gastric cancer patients. The most interesting result was the finding of a novel E-cadherin truncating germline mutation in exon 11, among the 15 (7%) early onset diffuse/mixed type gastric cancer patients. Interestingly, although no positive history of gastric carcinoma was reported for the family of this patient, the mother of the patient was affected with bilateral breast carcinoma at the age of 49 years. An association of breast carcinoma, in particular of the lobular type, with E-cadherin germline mutations was also found in our previous study,4 and a significant over-representation of this tumour type in families with E-cadherin germline mutations has been shown.25 In addition, somatic mutations of the E-cadherin gene are frequently found in sporadic, diffuse type gastric carcinomas and lobular breast carcinomas.26,27 Although no DNA of the patient’s mother was available to prove that she was a mutation carrier and the histopathological type of the breast carcinoma is unknown, it is intriguing to speculate that the bilateral breast carcinoma might have been associated with the E-cadherin germline mutation in this family, emphasising that breast carcinoma is part of the tumour spectrum associated with E-cadherin germline mutations. The finding of a somatic missense mutation in exon 10 in the tumour of the patient with the truncating germline mutation, which alters the LDFE-motif of a Ca-binding domain21 to LIFE, would predict a complete loss of normal cell adhesion and is consistent with the diffuse cytoplasmic protein staining pattern, which was demonstrated by immunohistochemistry. According to the classical inactivation theory of tumour suppressor genes, this missense mutation may represent the second hit that results in complete inactivation of the E-cadherin gene, although we did not prove that the germline and the missense mutation were located on different alleles.
In the familial gastric cancer group, one truncating mutation and one missense mutation were found among 30 (7%) diffuse/mixed type familial gastric cancer patients, and none was found in the five familial gastric cancer patients with an intestinal type of cancer. One of the germline mutations, previously reported by our group,4 was detected in the four families (25%) that met the strict criteria for HDGC. Overall, a high variability regarding the frequency of E-cadherin germline mutations has been reported in the literature. A low incidence (0–6%) was found in familial gastric cancer patients from Japan and Finland,7,28,29 and two missense mutations were identified among five (40%) patients with a familial aggregation of the disease from Korea.5 In contrast, E-cadherin germline mutations have been reported in 25% to 36% of diffuse type gastric cancer families fulfilling the strict criteria of HDGC defined by the IGCLC, which requires histopathological confirmation not only of the index patient, but also of the other family members.19 The frequency of germline mutations we found in families within the criteria of HDGC are in line with those reported recently.8 High detection rates of E-cadherin germline mutations in up to 100% has been reported for the Maori families in New Zealand and in studies including a high proportion of families with multiple cases of very early onset gastric cancer.1,6 This suggests that besides histopathological confirmation in respect to the inclusion criteria, variations due to different ethnic origins and due to a specific selection of analysed families exist, which may explain at least in part the different findings concerning the frequency of E-cadherin germline mutations in the different studies.
Among the two mutations identified in the familial group, a novel missense mutation in exon 15 was found. This mutation replaces a prolin by an arginin in the cytoplasmic domain of the protein, and represents a non-conservative amino acid change. This replaced prolin is conserved between human, mouse, and chicken E-cadherin,21 and the alteration was not found among the 50 control participants. Moreover, in the functional analysis, this mutant was shown to disrupt the E-cadherin abilities to mediate cell adhesion and to suppress cell invasion. All these findings together strongly support a pathogenic role for this missense mutation.
With respect to the analysis of the TP53 gene, we identified one germline mutation among the 34 (3%) familial gastric cancer patients we analysed, namely a missense mutation in exon 8 leading to a non-conservative amino acid change from arginin to cystein at codon 283 (R283C). Exon 8 belongs to the highly conserved region of the TP53 gene, and R283 represents a residue involved in DNA contact.30 The R283C mutation has been reported as a somatic mutation in various cancer types, including gastric carcinoma, and the R283H but not the R283C has been reported as a germline mutation (TP53 data bank: http:TP53.curie.fr/). Germline mutations of the TP53 gene are known as the molecular genetic cause of the Li-Fraumeni syndrome.31 The Li-Fraumeni syndrome is a complex, autosomally inherited cancer predisposition syndrome which is mainly characterised by the occurrence of multiple tumours in children or young adults, a predominance of soft tissue sarcomas, osteosarcomas, and breast cancer, and an excess of brain tumours, leukaemia, and adrenocortical carcinomas. Gastrointestinal tumours have been shown to be only occasionally associated with TP53 germline mutations,32 and only two families with a predominant occurrence of gastric carcinomas in several family members have been reported in the literature.11,14 No TP53 germline mutations were found in two studies from Japan analysing seven and 11 familial gastric cancer patients, respectively.33,34 In the family of the patient with the TP53 germline mutation in our study, gastric carcinoma was the predominant tumour type, with three affected family members. This does not point at the first glance to an association with the Li-Fraumeni syndrome in terms of its classical definition. Besides gastric carcinoma in this family, one family member died due to carcinoma of the liver at the age of 34 years, and another due to leukaemia at the age of 18 years, which represents a neoplasia originally identified as a component of the Li-Fraumeni-syndrome.
The patient with the TP53 germline mutation in our study was also a carrier of a rare missense variant (Ala592Thr) in the E-cadherin gene. This was found in a similar frequency in healthy control individuals in a previous study35 and in our present study, thus classifying this variant as a polymorphism, not casually related to HDGC. Moreover, in our in vitro system, cells expressing the A592T E-cadherin variant behaved as cells expressing the wild type protein, thus propounding its non-pathogenic nature. Interestingly, H. pylori infection was visible in the gastric carcinoma of the patient with the TP53 germline mutation. It has been suggested that H. pylori infection and TP53 act synergistically in gastric carcinogenesis36 and may be related to the development of gastric cancer in this patient.
No germline mutations were identified in the RUNX3 and HPP1 genes. The RUNX3 gene has been implicated recently as a tumour suppressor gene in gastric cancer.16 Inactivation by promoter hypermethylation and hemizygous deletion has been demonstrated in 40% of early and in 90% of advanced gastric cancers, and a missense mutation in the highly conserved RUNT domain has been found in one tumour. Furthermore, RUNX3 knockout mice developed hyperplasia of the epithelial cells of the stomach glands.16 In addition, RUNX3 is an integral component of the TGF-β induced signalling cascade, and it is well known that the TGF-β mediated cell signalling plays an important part in gastric carcinogenesis.37 Therefore, RUNX3 is an attractive possibility as a gastric cancer susceptibility gene. However, our finding of no germline mutations in familial diffuse and intestinal type gastric cancer families argues against a major role for RUNX3 in gastric cancer predisposition.
The HPP1 gene encodes for a transmembrane protein, which may have several roles in cell growth, maturation, and adhesion. A high frequency of inactivation of the HPP1 gene by promoter hypermethylation has been shown for hyperplastic polyps of the colon, and also for colorectal adenomas and colorectal and gastric carcinomas.38 In our study of 34 German familial gastric cancer patients and two families (one Canadian and one Portuguese) with a gastric polyposis phenotype, no HPP1 germline mutations were found. Although we may possibly have missed mutations by the DHPLC screening or larger genomic deletions, which are not detectable by the methods used, our results suggest that HPP1 and RUNX3 are not important alternative gastric cancer predisposition genes.
In conclusion, we report a frequency of 7% of E-cadherin germline mutations in a group of familial diffuse and mixed type gastric cancer patients from Germany. Our finding of a truncating germline mutation in an early onset diffuse type gastric cancer patient, whose mother was affected with bilateral breast carcinomas, underlines the association of E-cadherin germline mutations with this tumour type. The identification of a germline mutation in the TP53 gene suggests the TP53 gene should be included in genetic screening of familial gastric cancer patients, and supports a heterogeneous genetic predisposition to gastric cancer. In order to elucidate such predisposition, we have ruled out the RUNX3 and HPP1 as major gastric cancer predisposition genes.
Acknowledgments
We thank K F Becker and J Müller for critical reading of the manuscript.
REFERENCES
Footnotes
-
The study was supported by the German Cancer Foundation, Mildred-Scheel-Stiftung, grant 70-3124-Ke1.
-
Conflict of interest: none declared