Article Text

Statistics from Altmetric.com
Birt-Hogg-Dubé (BHD) syndrome, an inherited autosomal genodermatosis characterised by benign tumours of the hair follicle, is associated with renal neoplasia, lung cysts, and spontaneous pneumothorax.1 The novel causative gene, identified by linkage analysis in BHD families, is localised on chromosome 17p11.2.2 Protein truncating germinal mutations within a hypermutable (C)8 tract occur in patients with BHD syndrome and lead to an increased risk of kidney cancer.3
Microsatellite repeats are widely distributed throughout the genome. Owing to a defect in the DNA mismatch repair gene, a subset of tumours accumulates frequent deletion and insertion mutations in these repetitive DNA sequences.4–6 Most microsatellite instability (MSI) has so far been described in non-coding DNA within introns of intergenic regions in the genome. However, in some cancer related genes and mismatch repair genes, MSI has been identified in protein coding regions. The first target sequence identified within a coding region was a poly (A)10 nucleotide tract of the TGFBR2 gene.7 The other mutational targets of MSI have been found in repetitive sequences of IGF2R8 and BAX9 genes involved in the regulation of cell growth and in the promotion of apoptosis, respectively. Furthermore, frameshift mutations in repeat sequences of the DNA mismatch repair genes MSH3 and MSH6 have been reported.10 In tumours with MSI, the mechanism of tumorigenesis is believed to involve frameshift mutations of microsatellite repeats within the coding regions of genes, the inactivation of which is considered to contribute to tumorigenesis.
Early reports suggested that BHD syndrome was associated with a predisposition to colon neoplasms,11–13 but Zbar et al14 reported that colon cancer and colon polyps are not related to BHD syndrome. Recently, Khoo et al15 reported that colorectal neoplasia is an associated feature of BHD in some families. To elucidate whether the BHD gene is associated with colon neoplasia and one of the MSI target genes, we screened the poly (C)8 tract of the BHD gene, a mutational hot spot, in 32 MSI sporadic colorectal carcinomas and 80 microsatellite stable (MSS) sporadic colorectal carcinomas. In addition, we screened the entire coding region of the BHD gene in 13 MSI colorectal carcinoma cell lines and nine MSS colorectal carcinoma cell lines.
MATERIALS AND METHODS
Samples
As described in our previous study,16 we collected 325 consecutive patients who had undergone operation for primary colorectal cancers at Seoul National University Hospital, Seoul, Korea. Fifteen cases of familial colorectal cancers (two HNPCC, four FAP, and nine with a family history of cancer) were excluded through detailed family history taking, obtained by interviewing the patients and their relatives. Twenty-two colorectal cancer cell lines were obtained from the Korean Cell Line Bank, Seoul, Korea. Thirteen of these cell lines (SNU-175, SNU-407, SNU-769A, SNU-769B, SNU-1040, SNU-1047, SNU-C2A, SNU-C4, HCT-116, LoVo, LS174T, HCT-8, and DLD-1) are MMR negative and nine (SNU-61, SNU-81, SNU-283, SNU-503, SNU-1033, SNU-1197, Colo205, SW480, and WiDr) are MMR positive in terms of their MSI status, as determined by a previous study.17–19
Key points
-
Birt-Hogg-Dubé (BHD) syndrome, an inherited autosomal genodermatosis characterised by benign tumours of the hair follicle, is associated with renal neoplasia, lung cysts, and spontaneous pneumothorax.
-
Recently, the BHD gene containing a potential MSI target sequence has been identified as a cancer causing gene, which encodes a novel protein called folliculin.
-
Although earlier studies described the relation between colonic neoplasia and BHD syndrome, it was questionable that the BHD gene was associated with colon neoplasia.
-
We screened the poly (C)8 tract of the BHD gene, a mutational hot spot, and found mutations in five (16%) of 32 MSI sporadic colorectal carcinomas and in one (7.7%) of 13 MSI colorectal carcinoma cell lines. However, we were unable to find any frameshift mutation in 80 MSS sporadic carcinomas or nine MSS colorectal carcinoma cell lines. In addition, we found two heterozygous missense mutations, Arg137Cys and Arg462Ser, in SNU-1040 and LoVo cell lines with MSI, respectively.
-
The frequency (16%) of mutations in the BHD gene was the same as that in the IGF2R gene and this was less than that of the TGFBR2, MSH3, BAX, and MSH6 genes. Interestingly, all tumours with the BHD gene mutation harboured concurrent mutations of the poly (C)8 tract of the MSH6 gene (p=0.002), but the frameshift mutations of the BHD and IGF2R genes were mutually exclusive.
-
Our findings strongly support that the BHD gene is associated with colon cancer and putative MSI target genes involved in the development of MSI colorectal carcinomas.
DNA isolation and MSI analysis
In a total of 310 cases of sporadic colorectal cancers, genomic DNA from the tumour and corresponding normal tissues were procured from formalin fixed and paraffin embedded tissue samples by microdissection. MSI analysis of tumours was performed using an ABI 377 automatic sequencer (Perkin-Elmer, Foster City, CA) with fluorescent dye labelled primers of BAT-26 and BAT-25 markers. Thirty-two of 310 (10.3%) sporadic colorectal carcinomas were defined as MSI.
Mutation analysis of the BHD gene
To screen for mutations in the BHD gene, entire exons of the gene were examined by DNA sequence analysis. The primer sequences and the detailed reaction conditions for amplification have been described previously.3 Bidirectional sequences of PCR products were analysed using an ABI Prism 3100 Genetic Analyzer (Applied Biosystems, Foster City, CA).
Statistical analysis
To compare the frequencies of frameshift mutations of MSI target genes, we performed the χ2 test or Fisher’s exact test using SPSS software (version 10.05, SPSS Inc, Illinois, USA). Odds ratio (OR) and 95% confidence intervals (CI) were also calculated. A p level of <0.05 was considered statistically significant.
RESULTS
Three different mutations, a 1 bp and 2 bp deletion and a 1 bp insertion in the (C)8 repeat of the coding region of the BHD gene, were observed in five (16%) of 32 sporadic colorectal carcinomas (fig 1). In addition, HCT-116 in the MSI cell lines had a 1 bp insertion mutation in the poly (C)8 tract (1 of 13, 7.7%). As controls, none of 80 MSS sporadic colorectal carcinomas or the nine MSS colorectal carcinoma cell lines had mutations in the (C)8 repeat of the coding region of the BHD gene. The overall mutational profiles of the six genes evaluated showed diverse combinations (table 1). Among the 32 MSI tumours, 31 (97%) tumours had mutations in more than one gene, 25 (78%) tumours had mutations in more than two genes, 15 (47%) tumours had mutations in more than three genes, and five (16%) tumours had mutations in more than four genes. There were no homozygous mutations in the BHD, IGF2R, or MSH3 genes. Homozygous mutations in the BAX and MSH6 genes were rare, only one of 16 BAX mutations and two of 11 MSH6 mutation, whereas most of the TGFBR2 mutations were homozygous (13 out of 26, 50%). The frequency (16%) of mutations in the BHD gene was the same as that in the IGF2R gene and this was less than that of the TGFBR2, MSH3, BAX, and MSH6 genes. Interestingly, although the expected ratio of concomitant BHD-MSH6 mutation was only 5.4%, all tumours with the BHD gene mutation harboured concurrent mutations of the poly (C)8 tract of the MSH6 gene (p=0.002, OR =1.833, 95% confidence intervals 1.069 to −3.144), but the frameshift mutations of the BHD and IGF2R genes were mutually exclusive. Like tumours with a BHD frameshift mutation, HCT-116 harboured concurrent mutation of MSH6 and not IGF2R.
Mutation profile of the MSI target genes in 32 MSI positive sporadic colorectal carcinomas
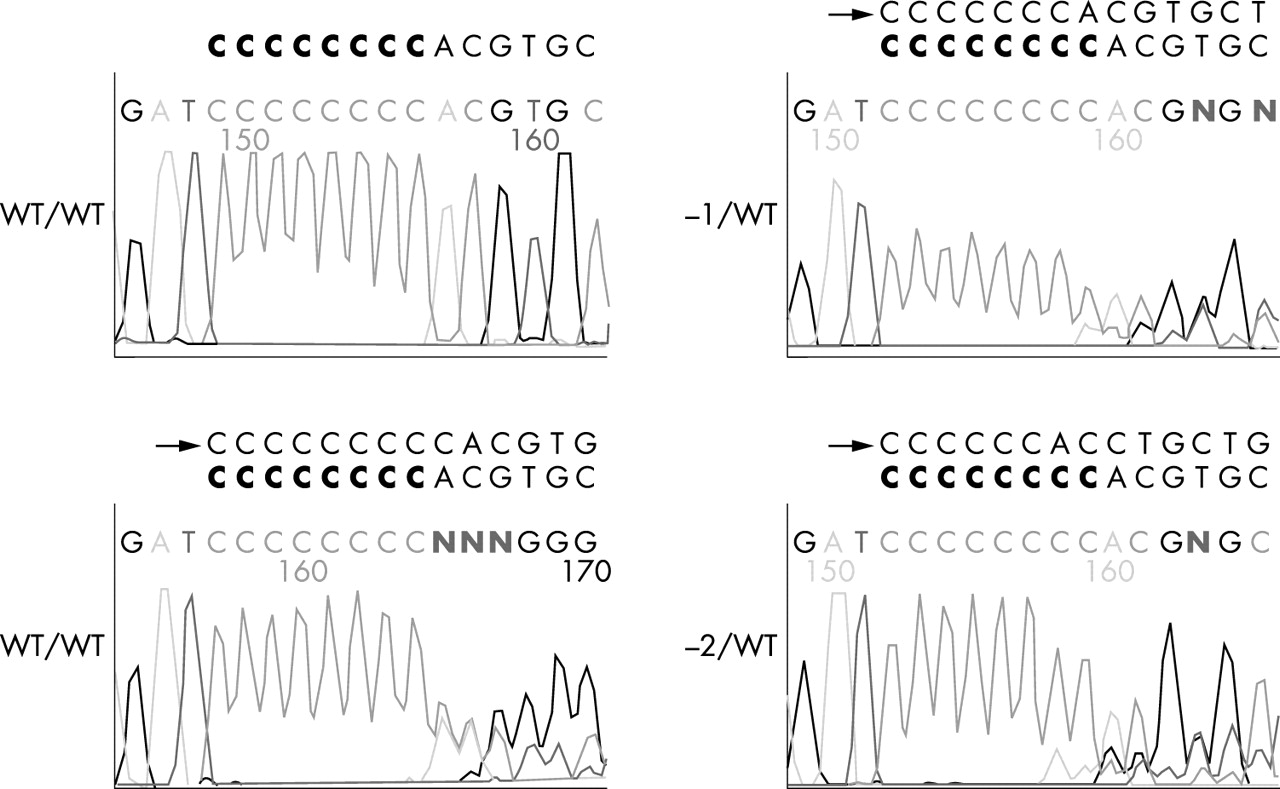
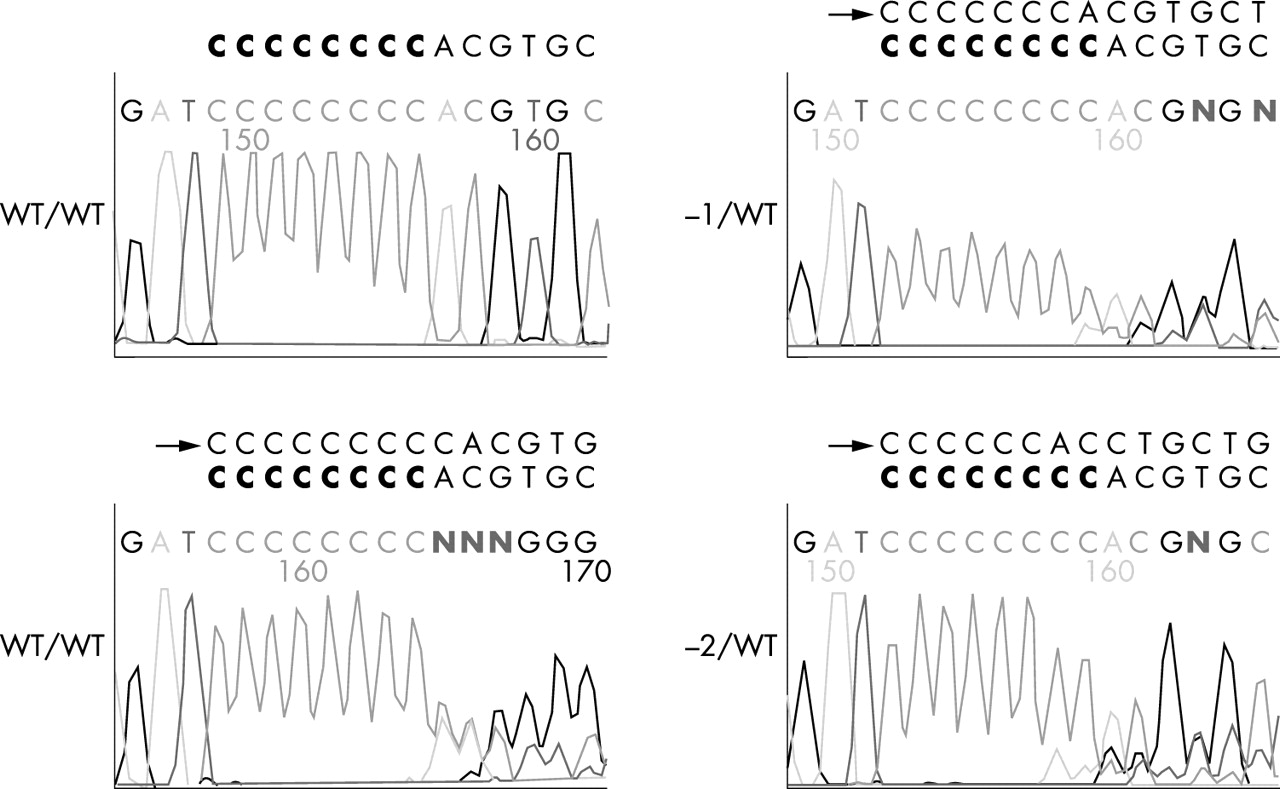{kind=link}
Sequencing analysis of the representative frameshift mutations of the BHD gene in MSI positive sporadic colorectal carcinoma. Arrows indicate insertion/deletion sequences in mononucleotide repeats. The wild type poly (C)8 tract is marked in bold letters.
To investigate further whether other types of mutation were found in both MSI and MSS colorectal cancer cell lines, we screened the entire coding region of BHD in 13 MSI cell lines and nine MSS cell lines. Two heterozygous missense mutations, Arg137Cys and Arg462Ser, were found in SNU-1040 and LoVo cell lines with MSI, respectively. However, no biallelic mutation was found in 13 MSI and nine MSS cell lines.
DISCUSSION
BHD syndrome is a rare inherited genodermatosis characterised by hair follicle hamartomas, kidney tumours, and spontaneous pneumothorax. Protein truncating mutations have been found in the BHD gene in BHD families, and most reported insertion/deletion mutations have been found within the poly (C)8 tract.3,15 These results suggest that the polycytosine mononucleotide tract is hypermutable and particularly prone to disease causing mutations. These insertion/deletion mutations resulted in a stop codon predicted to truncate the protein 26 missense amino acids downstream and 38 missense amino acids downstream, respectively. Truncated proteins lose the N-glycosylation site and the myristoylation site.3 In addition, we found two heterozygous missense mutations, Arg137Cys and Arg462Ser, in SNU-1040 and LoVo cell lines with MSI, respectively. These missense mutations result in an amino acid change from positively charged to uncharged amino acid. In addition, BHD protein, folliculin, was highly conserved among mouse, D melanogaster, and C elegans and this novel protein may have an important biological function in a wide range of organisms. Clues to the mechanism of tumorigenesis in sporadic colorectal carcinoma with a defective BHD gene may come from consideration of the BHD syndrome phenotype. Phenotypic data1 suggest that defective folliculin may affect the structure of the cytoskeletal network and the interaction between epithelial and mesenchymal cells. Perhaps the mutation alters the composition of the extracellular matrix, and produces a structural or microenvironmental abnormality that affects the regulation of cellular proliferation.
In most published studies of potential target genes, the prevalence of SMT (short mononucleotide tract) mutations in control, presumably unselected genes has been noted to be low. The study of such control genes has in general indicated that A or G tracts of 9 bp or fewer are mutated in <5% of cancers with MSI. Duval et al20 proposed an experimental cut off frequency value (10–15%) that divides mutational events into those that are or are not selected for during MSI-H tumour progression. In a recent paper,21 although real target genes were confirmed by comparing the frequency of mutation in the coding repeat with the non-coding repeat, these results were dependent on different comparative statistical approaches and showed the difference of significance. The mutation frequency is still likely to be a useful indicator of the functional role played by a specific gene. According to our results that 16% of MSI tumours harboured mutations of the poly (C)8 tract in the BHD gene, there is still reason for concluding that the BHD gene is a putative MSI target gene. To confirm that the BHD gene is a target of MSI colorectal cancer, general cytological approaches using gene array technologies are needed.
In summary, we found that mutations of the poly (C)8 tract of the BHD gene in MSI sporadic colorectal carcinomas and the frequency of mutations in the BHD gene is comparable with that of the IGF2R gene, which is one of the well known MSI target genes. In addition, two heterozygous missense mutations, Arg137Cys and Arg462Ser, were found in SNU-1040 and LoVo cell lines with MSI, respectively. Functional studies are required to understand the biological role of folliculin in colorectal carcinogenesis. Our findings suggest that the BHD gene is associated with colon cancer and putative MSI target genes involved in the development of MSI colorectal carcinomas.
Acknowledgments
This work was supported by the National Cancer Centre Research Grant (Research Grant 2001) and the 2002 BK21 project for Medicine, Dentistry, and Pharmacy.