Article Text

Statistics from Altmetric.com
- Pgp, P glycoprotein
- CRC, colorectal cancer
- CIN, chromosome instability
- HNPCC, hereditary non-polyposis colorectal cancer
- MSI, microsatellite instability
- MMR, mismatch repair
- MSS, microsatellite stable
- SSCA, single strand conformation analysis
- HA heteroduplex analysis
- DSCA, double strand conformation analysis
Pglycoprotein (Pgp), encoded by the MDR1 gene, is a transmembrane transporter that acts as an efflux pump in an ATP dependent fashion.1 Multidrug resistance, the main problem in efficient cancer chemotherapy, is mainly caused by increased expression and acquired mutations in the MDR1 gene.2 Pgp is expressed physiologically in epithelial cells of the kidney, liver, pancreas, and colon, suggesting its role in secretion of toxic compounds.3 Pgp is also expressed in the blood-brain barrier, adrenal glands, and lymphocytes where its role is still uncertain. Recently, additional functions for Pgp, including immune response4 and regulation of apoptosis,5 have been suggested in normal tissues and in cancers. High expression of Pgp at the apical surface of differentiated tubular structures was identified in previously untreated colorectal cancers (CRC)6 and its high expression at the leading edge of a colorectal carcinoma was associated with tumour progression.7
In contrast to the majority of CRC, which develop as a result of chromosomal instability (CIN), a proportion of sporadic CRC and 90% of cancers in patients with hereditary non-polyposis colorectal cancer (HNPCC) exhibit microsatellite instability (MSI).8 In MSI CRC, chromosomal aberrations such as large deletions, translocations, and gene amplifications are rare; the great majority of MSI tumours are usually diploid or near diploid. In MSI CRC, inactivation of the mismatch repair (MMR) system owing to mutations or methylation of MMR genes results in a 1000-fold accumulation of point mutations in oncogenes and tumour suppressor genes which trigger tumour progression. Therefore, in addition to genes mutated in microsatellite stable (MSS) CRC, such as APC, p53, and K-ras, there are other genes important in the development of MSI cancers. Interestingly, MSI CRCs are also more often resistant to several chemotherapy drugs; the selection of cells for resistance to cisplatin can result in the loss of DNA mismatch repair, and loss of DNA mismatch repair in turn contributes to resistance to cisplatin.9 Recently, a transcription factor complex TCF4/β catenin responsive element was identified in the MDR1 promoter region pointing to a direct link between the MDR1 gene and the WNT signalling pathway, the most important pathway altered in colorectal cancers.10 To determine the role of the MDR1 gene in the initiation and progression of CRC, we systematically screened the complete coding and promoter region of the MDR1 gene for alterations in a large cohort of patients with previously untreated colorectal cancer and in a normal control population. In this study, we report naturally occurring functional germline and somatic mutations in the MDR1 gene in patients with microsatellite unstable CRC and correlation of MDR1 functional polymorphisms with increased lymphoid infiltration in tumours with and without MSI.
MATERIALS AND METHODS
Patients
Between 1996 and 2000, 400 newly diagnosed colorectal cancer (CRC) patients from clinics all over Slovenia participated in this study. None of these patients had received chemotherapy treatment before operation. Primary colorectal adenocarcinomas as well as corresponding normal colorectal mucosa taken from a site several centimetres distant from the tumour were used in the study. Tumours were histopathologically evaluated according to the classification of Jass et al.11 The lymphoid infiltration of the tumours was assessed semiquantitatively by two independent pathologists. Accordingly, tumours with well represented lymphocytes along the advancing margin of the tumour were scored as positive for lymphoid infiltration.
Control samples
To determine the potential pathogenicity of MDR1 alterations detected in our study, we also analysed DNA from 100 unrelated unaffected blood donors.
DNA isolation
Colorectal tumours and corresponding normal tissue samples were snap frozen in liquid nitrogen and stored at −70°C. DNA was isolated after tissue digestion using standard phenol/chloroform extraction and ethanol precipitation.
Analysis of microsatellite instability
Microsatellite instability (MSI) analysis using a “reference panel” of microsatellite markers was performed in 400 unselected primary colorectal cancers (CRC) as described in our previous study.12 Thirty-eight tumours were defined as high microsatellite instability tumours (MSI-H) and analysed for MDR1 mutations.
MDR1 mutational analysis
We designed primers based on known genomic DNA sequences (Genbank accession numbers AC002457 and AC005068) specifically to amplify all 28 exons and exon/intron boundaries as well as the promoter region of the MDR1 gene in separate PCR reactions. Primer sequences and optimised PCR conditions are available at uros.potocnik{at}mf.uni-lj.si. For mutational analysis of the MDR1 gene, we used non-isotopic conformation analysis and silver staining. The basic principle of this method is a combination of three analyses which are all based on changes in three dimensional DNA structures, that is, single strand conformation analysis (SSCA), heteroduplex analysis (HA), and double strand conformation analysis (DSCA). We conducted them simultaneously on the same thin polyacrylamide gel. With this method, more than 95% of point mutations could be identified.13 Sequencing was performed with the BigDye Terminator Cycle Sequencing Ready Reaction Kit and ABI 310 sequencer (Perkin Elmer Cetus, Norwalk, CT, USA).
Methylation analysis
Tumour DNA was treated with sodium bisulphite to selectively convert only unmethylated cytosines in CpG sites of the MDR1 promoter region to thymines as described previously.14 To determine the methylation status of the MDR1 promoter, sequencing of bisulphite modified DNA using previously reported primers was performed.15 For sequencing we used the BigDye Terminator Cycle Sequencing Ready Reaction Kit and ABI 310 sequencer (Perkin Elmer Cetus, Norwalk, CT, USA). We scored a sample as hypermethylated if signals for unconverted methylated cytosines were higher than those for thymines converted from unmethylated cytosines in the majority of CpG sites after sequencing.
Immunohistochemical analysis of P glycoprotein expression
Sections of formalin fixed and paraffin embedded tissue blocks of tumour and normal intestinal wall were used. In short, a mouse monoclonal antibody (JSB-1, Biogenex) at dilution 1:10 was applied after microwave antigen retrieval (citrate pH 6.0, 15 minutes, at 850 W). For detection of the antigen, the LSAB method, using DAKO Tech-mate stainer, and DAB were used. According to the intensity of the imunnohistochemical staining estimated by two independent pathologists, samples were divided into three groups of low, moderate, and high Pgp expression. Samples with an estimated intensity of more than 50% (fig 1A, I) and less than 50% (fig 1J) of the most intensively stained sample were scored as high and moderate Pgp expression, respectively. Samples with no distinctive membrane staining (fig 1B, J) were scored as low Pgp expression. For statistical comparisons, moderate and high Pgp expression was evaluated against low Ppg expression.

Immunohistochemical staining for P glycoprotein using JSB-1 monoclonal antibodies. We observed moderate to high (A) and in some samples low (B) Pgp staining in epithelial cells of normal colonic mucosa. High Pgp staining was present in lymphocytes of normal mucosa, in particular in those forming lymphoid follicles (C, D, E). Even higher intensity of Pgp staining was observed in tumours compared to normal mucosa (F). In some tumour samples, high Pgp staining was present in lymphocytes of the infiltrating tumour border (G). The intensity of Pgp staining in tumours correlated with grading of tumours, being most intense in well differentiated (H), moderate in moderately differentiated (I), and low in poorly differentiated tumours (J).
Statistical analysis
We used the χ2 test with software package SPSS to compare clinicopathological characteristics between colorectal tumours with and without MDR1 polymorphisms. We used the two sided Fisher exact test to compare MDR1 mutations and methylation status with MSI status of colorectal tumours. We also used the two sided Fisher exact test to compare Pgp expression in unselected tumours (controls), tumours with functional polymorphisms, and MSI-H tumours. In all tests, p values of less than 0.05 were considered to indicate statistical significance.
RESULTS
Mutations and polymorphisms in the MDRI gene
We identified 12 different germline and five different somatic alterations in the MDR1 gene in initial screening of 60 patients with primary untreated colorectal cancer (CRC). Thirty patients with tumours exhibiting high microsatellite instability (MSI-H) and 30 patients with microsatellite stable (MSS) tumours were included in this initial screening. We confirmed the origin of the alterations by comparing DNA from tumour and from corresponding normal tissue. For exons with detected alterations, the study was extended to up to 350 patients with colorectal cancer and 100 unaffected blood donors. All alterations were identified as aberrant gel migration patterns with conformational analysis and further confirmed and characterised by direct sequencing of both DNA strands (forward, reverse) (fig 2). To exclude possible random errors, PCR and sequencing reactions were repeated at least three times for each alteration identified.

{kind=link}
{kind=link}
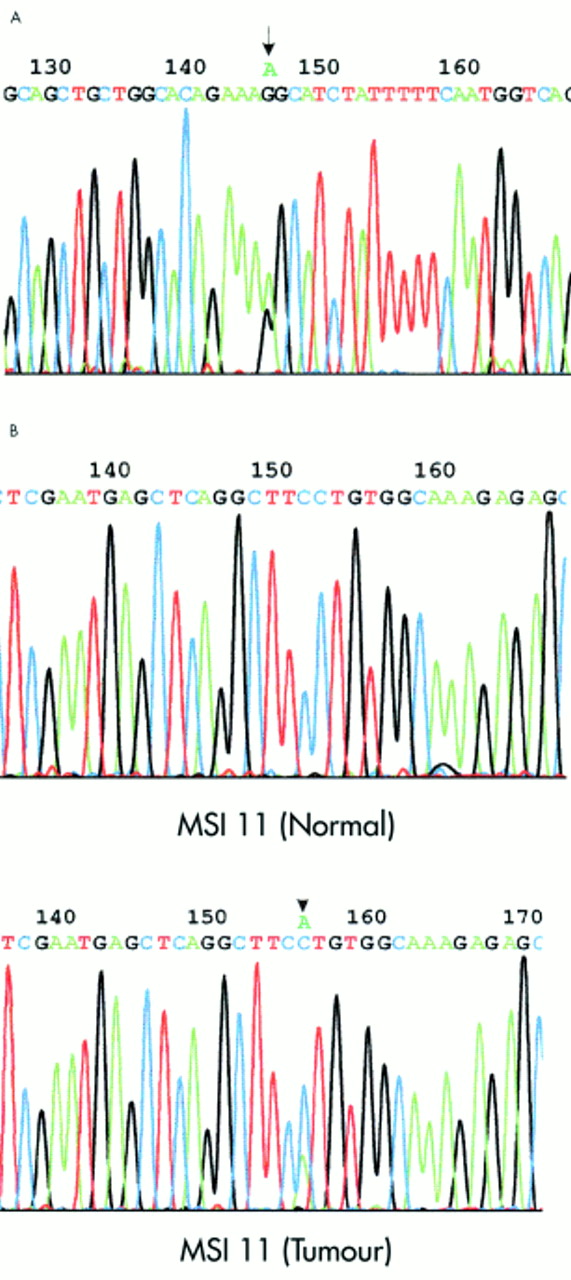
Sequence analysis of the MDR1 gene. (A) Germline mutation in exon 28 of the MDR1 gene identified in tumour sample MSI 11. Arrow indicates G>A substitution at position 3793 resulting in an amino acid change at position 1265 from non-polar glycine to polar serine (G1265S). (B) Somatic mutation in the MDR1 gene promoter region (−14 G>T) identified in tumour sample MSI 11. Arrow indicates C>A substitution (sequenced with reversed primer) 14 bp before the major transcription initiation site (numbering according to Kohno et al19 (MSI 11 (tumour)). Sequencing of the MDR1 promoter region in normal tissue from the same patient showed the somatic origin of this mutation (MSI 11 (normal)) .
Seven different MDR1 alterations, two germline and all five somatic, were only detected in 5/38 (13%) MSI-H tumours, but not in 400 patients with MSS tumours or in 100 controls (p<0.001, table 1). In 4/5 MSI-H tumours with unique alterations in the coding region there were concomitant alterations in the promoter region (p<0.001). A unique missense germline alteration and all somatic coding region alterations identified in MSI-H tumours were in the codons conserved throughout the species. On the other hand, all missense germline alterations (table 2) identified in MSS CRC patients and in controls were in codons not conserved throughout the species. The germline G1265S substitution identified in sample MSI 11 was of special interest (fig 2). It was the only germline substitution in the coding region that was found in the MSI-H tumour with a concomitant somatic promoter mutation (fig 2). G1265S was also the only germline alteration that resulted in substitution of an amino acid conserved through species; in this case non-polar Gly is replaced by polar Ser. The G1265S substitution is located in the predicted IC12 domain after the ATP binding site. It was not found on 200 chromosomes from normal blood donors or on 800 chromosomes from patients with CRC.
Mutations and germline promoter functional polymorphism in the MDR1 gene identified in colorectal cancers with microsatellite instability
Correlation between MDR1 polymorphisms and increased lymphoid infiltration in tumours
In contrast to two germline alterations that we found only in MSI-H tumours, the frequency of 10 other detected germline alterations (table 2) did not differ between MSI-H, MSS CRC, and controls. Comparison of more frequent MDR1 polymorphisms also identified in controls with clinicopathological data in patients with MSI-H and MSS CRC showed significant correlation between lymphoid infiltration in tumours and two polymorphisms, one in the promoter (+8 T>C, p=0.036) and the other in intron 1 (IVS1-81delG, p=0.010) (table 2). No significant correlation between MDR1 polymorphisms and age, sex, tumour location, tumour type, Duke's stage, and differentiation grade was observed.
Methylation analysis of MDR1 promoter
Genomic DNA from tumour and corresponding normal mucosa from 12 patients with MSI-H CRC and from 10 patients with microsatellite stable (MSS) CRC was analysed for MDR1 promoter methylation. We observed hypermethylation of CpG sites in the MDR1 promoter in tumour DNA from 10/12 (80%) of MSI-H tumours but in none of the 10 MSS tumours and corresponding normal mucosa (p<0.001). We observed hypermethylation of CpG sites in the whole MDR1 promoter region analysed rather than associated with specific CpG sites.
Pgp expression in colorectal tumours and corresponding normal mucosa
Colorectal tumours and corresponding normal mucosa from 14 colorectal cancer (CRC) patients with microsatellite stable tumours and without germline functional MDR1 polymorphisms (control group), 28 CRC patients with functional germline MDR1 polymorphisms, and 12 CRC patients with tumours exhibiting high microsatellite instability (MSI-H) were analysed for immunohistochemical Pgp expression (fig 1, table 3). For statistical comparisons, moderate and high Pgp expression was evaluated against low Ppg expression. We observed higher Pgp staining in cells from well and moderately differentiated tumours as compared to normal mucosa cells in all groups of CRC patients. Of 44 moderately and well differentiated tumours, 24 had high, 19 moderate, and one low Pgp expression while of 54 normal colon mucosa samples, 16 had high, 21 moderate, and 17 low Pgp expression (p=0005). Intensity of Pgp staining in tumours correlated with grading of tumours, being most intense in well differentiated (10 tumours high and one tumour moderate Pgp expression), moderate in moderately differentiated (14 tumours high, 18 tumours moderate, and one tumour low Pgp expression), and low in poorly differentiated tumours (10 tumours low Pgp expression) (p<0.0001). In less differentiated tumours, Pgp expression was limited to the inner surface of the glands. We observed lower Pgp expression in MSI-H tumours as compared to tumours in the control group (p=0.042). Although not statistically significant, Pgp expression tended to be lower in epithelial cells of normal colon mucosa in CRC patients with functional MDR1 polymorphisms and in CRC MSI-H patients as compared to the control group (table 3).
Immunohistochemical staining for P glycoprotein (Pgp)
DISCUSSION
In this study, we provide detailed and extensive analysis of germline and somatic alterations in the promoter and complete coding region of the MDR1 gene in patients with previously untreated colorectal cancer (CRC) and in a normal control population. We have identified 12 different germline and five different somatic alterations. Of 10 different germline alterations identified in patients and controls, five were in non-coding regions, two did not change any amino acids, and three changed non-conserved amino acids. Two unique germline and all five somatic mutations were identified only in patients with colorectal cancers exhibiting high microsatellite instability (MSI-H, table 1). To the best of our knowledge, this is the first report of naturally occurring somatic mutations in the MDR1 gene in cancer patients, as previously only acquired mutations were reported in resistant tumours after chemotherapy.2 Of the somatic coding mutations, there was one frameshift and two missense mutations changing amino acids conserved throughout species. Also, one unique germline mutation changed a conserved amino acid which results in changed polarity at this position (sample MSI 11, fig 2). Identified MDR1 mutations in MSI-H tumours might change the functional activity of Pgp. The functional testing of these mutations in transfected cells is under way. In 4/5 MSI-H CRC with coding MDR1 alterations, we identified concomitant alterations in the promoter region (p<0.001). Two MSI-H CRC (samples MSI 3 and MSI 5) had germline promoter mutation and somatic mutation in the coding region, sample MSI 11 had somatic mutation in the promoter and germline mutation in the coding region, and sample MSI 19 had two somatic mutations in the promoter and coding regions, respectively. Since it is unlikely that promoter and coding mutations would appear in the same tumours by chance, it is possible that MDR1 mutations in these MSI-H tumours are selected during tumorigenesis. Interestingly, none of these MSI-H tumours had mutations in the coding regions of major mismatch repair genes as determined in our previous study.12 Only 1/5 somatic MDR1 mutations identified in MSI-H CRC in our study was a deletion of one adenine in the (A)5 tract in exon 20 of the MDR1 gene (2422-2426 del A, sample MSI 19), although inactivating frameshift mutations resulting from 1 bp deletions or 1 bp insertions in mononucleotide tracts of genes are usually found in MSI-H CRC. The missense mutations in the coding region of MDR1 do not necessarily inactivate Pgp function, but could even increase Pgp functional transport activity2 or could alternatively alter the substrate spectrum.16 The high Pgp activity might protect intestinal cells against a wide variety of caspase dependent death stimuli, including FasL, tumour necrosis factor, and UV radiation5 and therefore contribute to cancer progression. Pgp might be a downstream target of WNT signalling pathway, the most important pathway altered in colorectal cancers, as a transcription factor complex TCF4/β catenin responsive element was identified in the MDR1 promoter region.10 In addition to the role in tumour progression, MDR1 mutations might also contribute to resistance to several chemotherapeutic agents often associated with MSI cancers.
We identified two polymorphisms, one in the promoter (+8 T>C, p=0.036) and one in intron 1 (IVS1-81delG, p=0.010), associated with lymphoid infiltration in tumours from CRC patients with or without MSI (table 2). CRC patients with these two polymorphisms tended to have lower Pgp expression in epithelial cells (table 3). In particular, the germline polymorphism in the promoter region might have an important influence on Pgp functional activity. We found the same polymorphism in two MSI-H CRC with concomitant somatic mutations in the coding region (samples MSI 3, MSI 5, table 1). However, 12 CRC samples with promoter polymorphism (+8 T>C) but without MSI were included in mutation screening of the complete MDR1 coding region, but no additional mutations were found. Previously, this promoter polymorphism has been associated with haematological malignancies.17 Our results might support the previously suggested role for Pgp in immune response.4,18
-
In this study we provide a systematic analysis of naturally occurring germline and somatic alterations in the MDR1 gene coding for P glycoprotein (Pgp) ABC transporter. The promoter and complete coding region of the MDR1 gene was analysed in 400 patients with previously untreated colorectal cancer (CRC) and in a control normal population.
-
We identified 12 different germline and five different somatic alterations. Two unique germline and all five somatic alterations were identified only in CRC patients exhibiting high microsatellite instability (MSI-H ). In 4/5 MSI-H tumours with mutations in the coding region, there were concomitant mutations in the promoter region (p<0.001).
-
We observed low Pgp expression in half of the MSI-H tumours mainly associated with poorly differentiated MSI-H tumours. We observed a significant increase (>80%) in methylation of CpG sites in the MDR1 promoter in tumour DNA from 10/12 (80%) of MSI-H tumours.
-
We also identified two germline MDR1 polymorphisms associated with lymphoid infiltration, one in the promoter (+8 T>C, p=0.036) and one in intron 1 (IVS1-81delG, p=0,010) in both MSI-H and tumours without MSI. These two polymorphisms were associated with lower Pgp expression.
-
These results suggest that the MDR1 gene may be involved in initiation and progression of MSI-H tumours. Our results may also support the previously suggested role for Pgp in immune response.
Consistent with previous reports, we observed higher Pgp staining in tumour cells as compared to normal mucosa cells even in untreated tumours. Pgp expression was correlated with tumour differentiation. The lower Pgp expression we observed in MSI-H tumours might be associated with poor differentiation of MSI-H tumours and hypermethylation of the MDR1 promoter. We found hypermethylation of the MDR1 promoter in 10/12 (80%) MSI-H tumours, including 5/6 MSI-H tumours with MDR1 mutations, but in none of the 10 MSS tumours and corresponding normal mucosa (p<0.001). Two MSI-H CRC samples (MSI 14 and MSI 22) that lack MDR1 hypermethylation also lack hMLH1 hypermethylation as we determined in our previous study.12 The other 10 MSI-H CRC analysed had methylation in both genes MDR1 and hMLH1, suggesting the more general failure of correct methylating mechanism in these tumours.
In conclusion, we identified germline and somatic mutations in the MDR1 gene associated with MSI-H tumours suggesting a role of MDR1 in progression of at least a subgroup of MSI-H tumours. Finding of novel genes mutated in MSI-H CRC might help in the understanding of tumorigenesis and to design appropriate treatment. The characterisation of naturally occurring functional polymorphisms and mutations in the MDR1 gene could also provide a good basis for functional studies of Pgp.
Acknowledgments
The Ministry of Education, Science, and Sport of the Republic of Slovenia supported this study (Project J3-7919-0381). The scope of the study was agreed by the Medical Ethical Commission of the Republic of Slovenia in October 1995. We are grateful to patients who participated in the study. We thank Rastko Golouh and Stanislav Repše for patient tissue samples. We thank Anton Cerar for help with the evaluation of imunnohistochemistry results. We also thank Teo Zizek and Aleš Novak for excellent technical assistance and Matej Bracko for statistical calculations.