Article Text

Statistics from Altmetric.com
Editor—An imbalance of genetic material, especially monosomy, will usually give rise to an abnormal phenotype. A few instances of proximal 2q deletions have been published, but previous cases (q12-q14,1 q12-q14.2,2q14-q213 4) have been associated with clinical features such as mental retardation, facial dysmorphism, heart defects, and renal and digital anomalies.3 We ascertained an interstitial deletion of chromosome 2 at q13-q14.1 (fig 1) in a clinically normal G6, P2, SAB3 woman aged 38. She had been referred for chromosome analysis following three successive miscarriages at 8½, before 11, and at 7 weeks' gestation. Her current pregnancy was chromosomally normal at amniocentesis and continuing at 26 weeks. Testing of her two previous children was not being pursued at the time of writing. This deletion was subsequently found to have been transmitted by her G2, P2 mother who had no associated phenotype nor history of miscarriages. Cytogenetic analysis at the 600 band level failed to detect a subtle insertional translocation of the missing material elsewhere in the genome and the partners of both mother and daughter had normal karyotypes. Here we have attempted to define the extent of this deletion and thereby the degree of euchromatic loss using fluorescence in situ hybridisation (FISH).
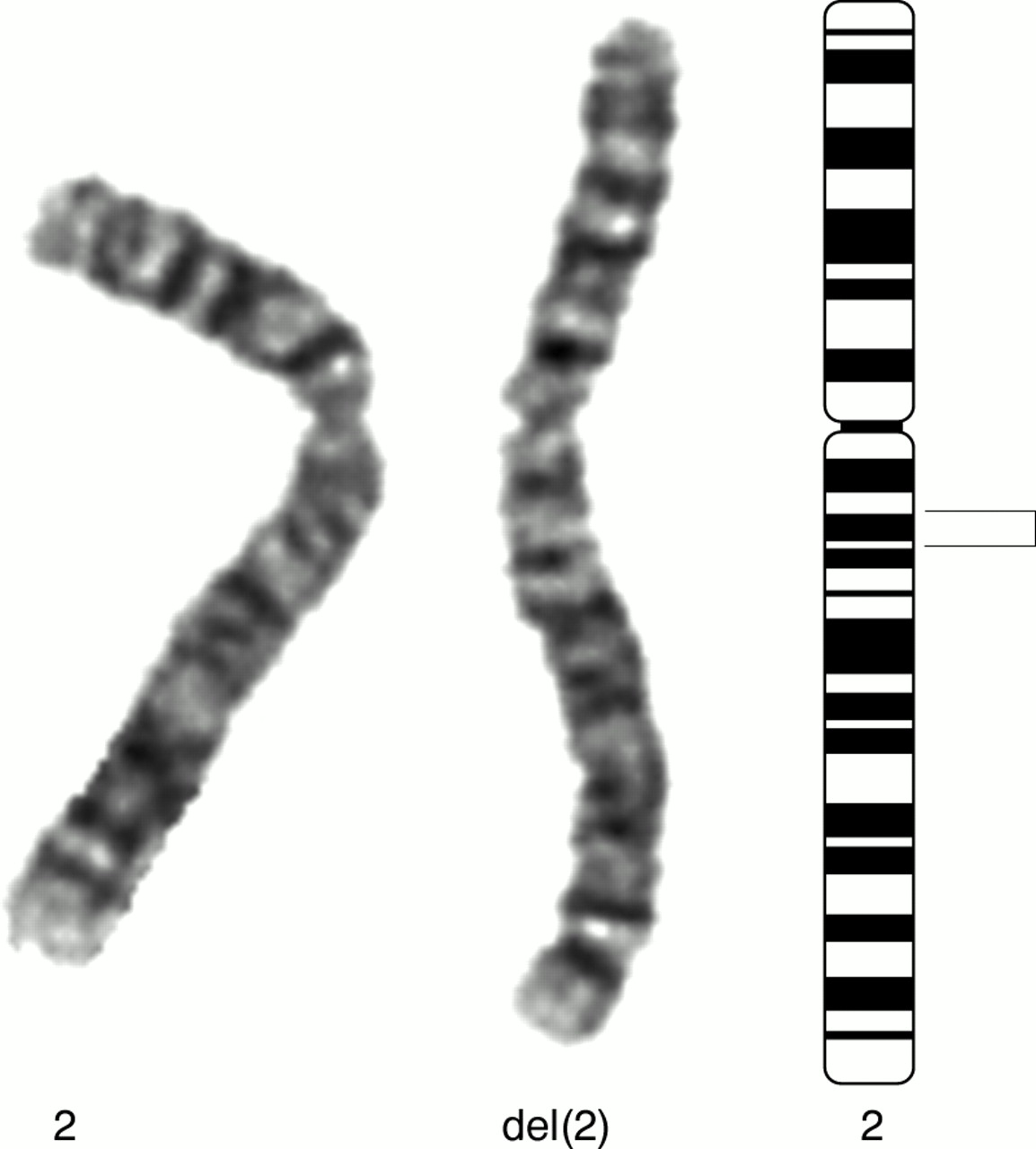
Chromosome 2 with deletion indicated by bracket. G banded partial karyotype (left) and ideogram (right).
Whole chromosome 2 paint (Cambio) corroborated our cytogenetic results in that no fluorescent signal was detected outside the two chromosomes 2 to account for the missing bands at 2q13-q14.1. To exclude an intrachromosomal insertion and to help identify the loci most proximal and distal to the deletion, a range of eight YACs (YAC Screening Centre, Milan) spanning proximal 2q were applied (table 1). At least five metaphases in either the proband or her mother were scored for a positive signal at each of these loci along with control probes at either the centromere (D2Z1) or distal to the deletion at 2q21 (845f5).
Details of YAC for FISH
Results showed that both proximal 2q14 YACs 786a12 and 817b4 were deleted from the abnormal chromosome 2 (fig 2), while the distal 2q14 YACs 679d2 and 821h9 were retained. We conclude, therefore, that consistent with our findings from G banded analysis, the proximal region of 2q14 is absent. Hybridisation with the 2q13 YAC 791f4 produced a signal on both homologues, but one signal was frequently reduced in size. To confirm a partial deletion of this YAC, we carried out dual colour FISH in which YAC 791f4 was detected using texas red and one of the known deleted YACs with fluorescein (786a12). This showed that the reduced signal was consistently associated with the deleted homologue (fig 3). Semiquantitative FISH5 in a series of 10 cells showed that the signal from the partially deleted YAC was only 0.48 times (95% CI 0.32-0.72) that of the signal from the normal homologue. All other YACs hybridised to both homologues without significant differential signal strength. Thus, this deletion extends from a proximal 2q13 breakpoint within YAC 791f4 to a distal 2q14.1 breakpoint within the 5 cM interval between YACs 786a12 and 679d2. The karyotype of the daughter can be summarised as 46,XX,del(2)(q13q14.1)mat.ish del(2)(q13q14.1) (D2Z1+, D2S135+, D2S1895 dim, D2S308-, D2S363-, D2S110+, D2S347+, D2S1888+, D2S112+, wcp2+).

FISH metaphase with YAC 786a12 (red signal) and the alphoid chromosome 2 centromere probe (green signals). Hybridisation was absent on the deleted chromosome, as indicated by the white arrow.

{kind=link}
{kind=link}
{kind=link}
FISH metaphase with YACs 791f4 (red signals) and 786a12 (green signals). 786a12 is absent and 791f4 signal strength reduced on the deleted chromosome, as indicated by the white arrow.
In this family, therefore, a deletion of 2q13-q14.1 confers no phenotypic effect despite spanning a region of approximately 4.5-6.0 Mb (estimated from a haploid autosomal length of 0.15-0.20%). To date, asymptomatic transmitted deletions of at least eight distinct regions of the human genome have been reported6-13 and this family provides a ninth example. Several theories have been postulated to account for the lack of phenotypic consequences. These include the late replication,6 7 9 12 low gene density,14 15 or non-essential DNA content of G dark bands,6 7 9 14 16 and haplosufficiency or dosage compensation of hemizygous loci.6 7 9 14 16 In our case, the size of the deletion is consistent with partial loss of G light 2q13 proximal to G dark 2q14.1 and, therefore, it is unlikely that the deleted material consists of entirely non-essential, non-coding DNA. On the other hand, it is known that 2q12-14 is not gene rich.15 Indeed, Saccone et al 15 describe R bands, with the exception of telomeres, as generally containing a moderate/low gene density (only 9% of R bands have a high gene content). In accordance with this, very few known genes lie within 2q13-q14. The inhibin beta chain precursor and interleukin-1 receptor antagonist protein precursor genes are located somewhere between 129 and 134 cM from the p telomere of chromosome 2,4 17 which roughly corresponds to the maximum extent of the deletion from 128 cM (YAC 791f4) to 135 cM (YAC 676d2).
Evidence that this region contains material of genetic significance is suggested by a cytogenetic case previously ascertained in this laboratory; an apparently identical, but de novo, partial deletion of band 2q14.1 was found in a girl of 16 referred with developmental delay, a cleft palate, and facial features suggestive of 22q11 deletion syndrome. A 22q11 deletion was excluded by FISH. There are at least three possible explanations for the contrast between the de novo and transmitted deletions. First, the deletion could be coincidental to the phenotype in the de novo case. Second, the phenotype associated with deletions of 2q13 to 2q14 may extend into the normal range. Third, these deletions may be cytogenetically identical but differ in breakpoint location and extent at the submicroscopic level. Unfortunately, no further material has been available with which to test the extent of the deletion in the de novo case with the same set of probes used in this report. Consequently, it has not been possible to explore further why a deletion associated with deleterious effects in one person is stably transmitted from one generation to the next in a family with no discernible phenotypic consequences.
Acknowledgments
We should like to thank Heather Reed for referring this family and providing additional information on their obstetric history. We also acknowledge John Crolla for his help with this project, Sarah Beal for preparing YACs and control probes for FISH, Nikki Savage for analysis of the de novo 2q deletion, Christine Joyce and Andrew Fisher for their help in the preparation of this report, Mark Redford for his assistance with the illustrations, and Judy Gladding for preparing the manuscript.