Article Text

Abstract
Clinical otosclerosis (OMIM 166800/605727) has a prevalence of 0.2-1% among white adults, making it the single most common cause of hearing impairment in this group. It is caused by abnormal bone homeostasis of the otic capsule with the consequent development of sclerotic foci that invade the stapedio-vestibular joint (oval window) interfering with free motion of the stapes. Impaired ossicular chain mobility results in a conductive hearing loss.
We identified the first locus for otosclerosis (OTSC1) on chromosome 15 in 1998 and reported a second locus (OTSC2) on chromosome 7 last year. Here we present results of a genome wide linkage study on a large Cypriot family segregating otosclerosis. Results of this study exclude linkage to OTSC1 and OTSC2 and identify a third locus, OTSC3, on chromosome 6p. The defined OTSC3 interval covers the HLA region, consistent with reported associations between HLA-A/HLA-B antigens and otosclerosis.
- otosclerosis
- linkage analysis
- HLA region
- chromosome 6
Statistics from Altmetric.com
Clinical otosclerosis (OMIM 166800/605727) has a prevalence of 0.2-1% among white adults, making it the single most common cause of hearing impairment in this group.1,2 The disease is characterised by isolated endochondral bone sclerosis of the labyrinth capsule that invades the stapedio-vestibular joint (oval window), interferes with free motion of the stapes, and causes a conductive hearing loss.3 Mean age of onset is in the third decade and 90% of affected persons are under 50 years of age at the time of diagnosis.2,4 Long term follow up suggests that approximately 10% of persons with otosclerosis ultimately develop a profound sensorineural hearing loss across all frequencies.4,5
The aetiology of otosclerosis is not understood and environmental as well as genetic factors have been implicated. Several epidemiological studies have indicated that familial segregation is most consistent with autosomal dominant inheritance with reduced penetrance.6–11 Temporal bone studies document two forms of otosclerosis. The first, histological otosclerosis, occurs in about 2.5% of the white population.12 The observed otosclerotic foci do not impair stapedial movement and therefore remain clinically inconsequential. The second, clinical otosclerosis, has a lower prevalence (0.2-1%), interferes with ossicular chain mobility, and leads to conductive and/or mixed hearing loss.1–5
In 1998, we localised the first otosclerosis gene (OTSC1) to chromosome 15q25-26 and last year we reported a second locus (OTSC2) on chromosome 7q34-36.13,14 These loci were identified in large autosomal dominant otosclerosis pedigrees from India and Belgium, respectively. Recently, in a study of nine European families segregating otosclerosis, we showed that at least one additional locus must exist.15 In this report, we describe the localisation of OTSC3 to chromosome 6p21.3-22.3.
METHODS
Clinical data
The Cypriot family presented in this study segregates autosomal dominant otosclerosis (fig 1). After informed consent was obtained, health and hearing histories were taken and a physical examination was performed to exclude chronic ear disease. Audiological testing was performed on all subjects (air conduction at 250, 500, 1000, 2000, 4000, and 8000 Hz; bone conduction at 500, 1000, 2000, and 4000 Hz). Audiograms were analysed based on age and sex dependent audiometric curves according to the International Organization for Standardization (ISO) 7029 standards. Owing to the variable age of onset of otosclerosis, only family members who were at least 50 years old and had hearing thresholds better than age adjusted median values (ISO 7029 standards) were considered unaffected. Two relatives by marriage with normal hearing were also considered unaffected. All persons considered affected had surgically confirmed otosclerosis (5) or had hearing loss consistent with the clinical diagnosis of otosclerosis (4).

Pedigree used to localise OTSC3 to chromosome 6p21.3-22.3. The linked haplotype is shown in green (* subjects used in first pass screen, + subjects added for the second pass scan, # subjects added for the final screen; III.10 with putative incomplete penetrance).
Marker analysis
Genomic DNA was isolated from study participants from whole blood samples using QIAamp DNA blood maxi kit (QIAGEN Inc, USA). Purified DNA was analysed by polymerase chain reaction (PCR) amplification using standard techniques.13,14 Genetic markers from the Cooperative Human Linkage Center screening set (Research Genetics, Weber version set 8, 8A) covering the 22 autosomes were run approximately every 20 cM. Additional microsatellite markers from the Marshfield Medical Research Foundation genetic map were used to analyse specific regions of interest further. Haplotype construction was done by hand and confirmed using Cyrillic version 2.1.
Linkage analysis
The LODSCORE, MLINK, and LINKMAP subprograms of the FASTLINK computer program were used to calculate two point, pairwise, and multipoint lod scores for linkage between markers and the disease locus.16 In regions generating a multipoint lod score of 1.0 or greater, additional markers and subjects were typed. Recombination frequencies between males and females were assumed to be equal.
For each marker, the number of alleles in the lod score calculations was set at the observed number of alleles in the pedigree (N); allele frequencies were set at 1/N. The frequency of the disease allele was set at 0.0001, assuming autosomal dominant inheritance and 70% penetrance, and the phenocopy rate was set at 1%. Different linkage parameters were used in lod score calculations over candidate intervals to confirm linkage results.
RESULTS
First pass genome scan
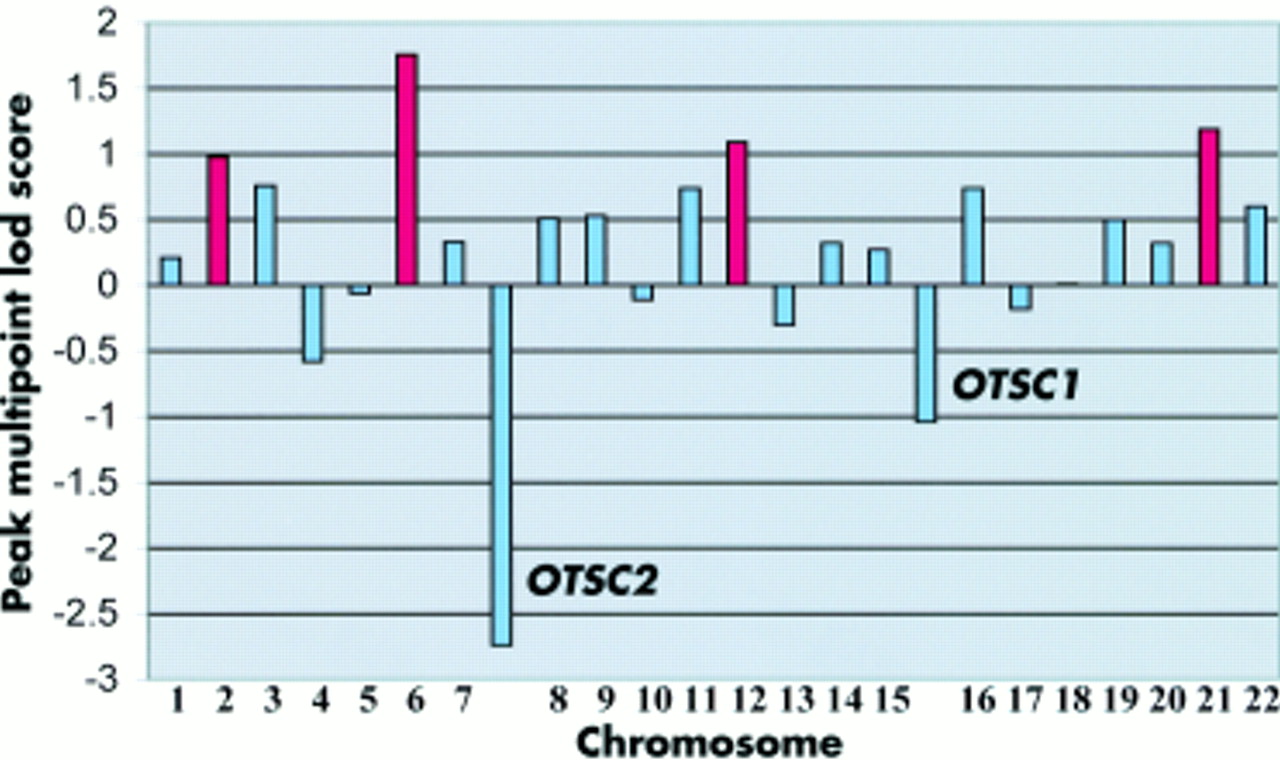
Linkage to the known otosclerosis loci on chromosomes 15q25-26 (OTSC1) and 7q34-36 (OTSC2) was investigated using published linked markers13,14 in seven affected persons and three unaffected persons from the Cypriot family (fig 1). Multipoint calculations were performed using four point rolling lods and generated maximum negative lod scores of −1.04 and −2.74 for OTSC1 and OTSC2, respectively. These results suggested that a novel otosclerosis gene was segregating in this family, and a genome wide screen was completed using 417 markers (fig 2).
{kind=link}
{kind=link}
Summary of peak multipoint lod scores by chromosome and OTSC1 and OTSC2 intervals generated after the first pass scan. In four regions (chromosomes 2, 6, 12, and 21), the peak multipoint lod score was approximately 1 or greater (shown in red).
First pass screening generated a multipoint lod score >1 over 1% of the genome. Nine percent of the genome had a lod score between 0 and 1, 31% a lod score between −1 and 0, 28% a lod score between −2 and −1, and 31% a lod score below −2. Table 1 summarises peak multipoint lod scores ≥1 and region sizes (lod>0) following the first pass screen. The highest lod score, 1.76, was detected on chromosome 6p21-22 (Zmax=1.76, 25 cM). Regions with suggestive linkage were also found on chromosomes 2q37 (Zmax =0.98, 18 cM), 12q24 (Zmax =1.09, 15 cM), and 21q22 (Zmax =1.19, 20 cM).
Lod scores generated over four candidate regions identified by the first pass screen
Second pass mapping of chromosomes 2q37, 6p21-22, 12q24, and 21q22
Second pass screening was completed over regions of chromosomes 2q37, 6p21-22, 12q24, and 21q22 (fig 2). In this screen, five markers were used from each candidate region to increase map density. Recalculation of the multipoint lod score on chromosome 21q22 generated a Zmax of 0.04. In the second pass screen of the remaining three regions, five additional family members were included and haplotyped. In two regions, chromosomes 2q37 and 12q24, negative peak multipoint lod scores of −1.34 and −2.55, respectively, were obtained. The peak multipoint lod score over the region of chromosome 6p21-22 increased to 3.06 (table 1).
OTSC3 maps to chromosome 6p21.3-22.3
The final screen included an additional seven persons belonging to the pedigree and examined by two of us (CAC, CG) during a recent trip to Cyprus. Seven more markers from the region also were added for two point lod score calculations, the maximum of which was 3.34 at D6S1660 with a recombination fraction (𝛉) of 0.001 (table 2). The maximum multipoint lod score was 3.83 assuming 70% penetrance, a phenocopy rate of 1%, and a disease gene frequency of 0.0001. Changing these parameters had only a minimal effect on the lod score except when penetrance was reduced to 40% (table 3). Even at this level, Zmax was 3.47, consistent with strong linkage between this region and the disease gene.
Two point lod scores for chromosomal region 6p21.2-22.3
Maximum chromosome 6 multipoint lod scores
Twelve tightly linked chromosome 6p markers were used to construct haplotypes (fig 1). Flanking markers were defined by subjects III.1, III.2, and III.8 (GAAT3A06, telomeric) and subjects II.1 and III.6 (D6S1680, centromeric). Therefore, the 17.4 cM region between GAAT3A06 and D6S1680 harbours an otosclerosis gene. III.10 (aged 52) appears to have inherited the affected haplotype but does not have a conductive deficit, a finding consistent with incomplete gene penetrance.
OTSC3 localises to an interval on chromosome 6p21.3-22.3 that includes the HLA region. The physical position of this interval extends from 23.7 Mb to 46.0 Mb based on the Ensembl map from EMBL - EBI and the Sanger Institute.
DISCUSSION
Although three otosclerosis loci have been mapped, no causative genes have been identified and we have little idea of the molecular processes involved in this disease. Temporal bone studies indicate that the first histological sign of incipient otosclerosis is resorption of bone around blood vessels with secondary enlargement of perivascular spaces.17 Immature bone rich in ground substance and deficient in collagen is laid down in these spaces and actively resorbed and remodelled. Gradually, mature bone with increased collagen and less ground substance is produced.17 Sites of predilection are the globuli interossei, embryonic rests of cells from the original cartilaginous framework of the inner ear.18
The apparent association of otosclerosis with cartilage has prompted many investigators to explore this relationship further. In a comparative study of antibodies to collagens type I, II, III, VI, IX, and XI, Bujía et al19 found significantly higher antibody levels to collagens type II and IX in persons with otosclerosis compared with unaffected controls. Further implicating collagens is an association study by McKenna et al18 in which a specific COL1A1 SNP allele was found more frequently in affected persons than controls.
In light of these finding, the localisation of OTSC3 to chromosome 6p is particularly interesting because the defined interval includes as candidate genes RING1 and COL11A2. Recent studies have shown that RING1 and Yin Yang 1 binding protein (RYBP) interact with Yin Yang 1 (YY1),20 a transcriptional activator of the COL1A1 promoter in fibroblasts.21 The regulatory elements of the COL1A1 promoter that respond to YY1 are different in humans and mice, noteworthy since mouse models of otosclerosis have not been reported. Abnormalities in the regulation of COL1A1 transcription could impact the normal stoichiometry of COL1A1 and COL1A2 production, as exemplified by osteogenesis imperfecta (OI), a disease also associated with fixation of the stapes footplate. Numerous mutations in COL1A1 and COL1A2 cause OI, including a mild phenotype (OI type I) most frequently resulting from abnormal steady state levels of COL1A1 mRNA. In an analysis of total cellular and nuclear RNA, Willing et al22 found that mutations predicting premature termination of COL1A1 reduce amounts of mRNA from the mutant allele and lead to a decrease in COL1A1. This change in protein level impacts type I collagen production, which is a heterotrimer of COL1A1 and COL1A2 in a 2:1 ratio. It is possible that mutations in RING1 impact COL1A1 transcription resulting in an OI type I phenotype so mild as to be limited to disease of the otic capsule.
COL11A2 is also an intriguing candidate gene. It is expressed in the bony labyrinth and causes autosomal dominant non-syndromic hearing loss at the DFNA13 locus.23 The promoter of COL11A2 includes a putative human specific collagen modulating element that acts cooperatively with G/CBE binding proteins to regulate procollagen transcription.
The localisation of OTSC3 to chromosome 6p is consistent with studies that have explored the relationship between HLA antigens and otosclerosis. Gregoriadis et al24 determined 27 HLA-A and B antigens in 68 Greek persons with surgically confirmed otosclerosis, and in comparison to 400 unrelated controls found a highly significant increase in HLA-A11, Bw35, and B14 in persons with a positive family history.24 HLA antigen biases with otosclerosis have also been reported in India25 but not in Sweden,26 suggesting that the frequencies of the different genetic forms of otosclerosis differ between populations. The analysis of additional Cypriot families segregating otosclerosis may expedite the cloning the OTSC3 gene.
Acknowledgments
The first two authors contributed equally to this work. We thank the families who made this research possible. This research was supported in part by NIH grant R01DC05218 (GVC and RJHS).
Electronic database information. The accession number and URL for data in this article are as follows: Center for Medical Genetics, http://research.marshfieldclinic.org/genetics/ (for marker order and genetic positions). Ensembl, http://www.ensembl.org/ (for chromosome bands and physical positions). Online Mendelian Inheritance in Man (OMIM), http://www.ncbi.nlm.nih.gov/Omim/ (for otosclerosis, MIM 166800/605727).