Article Text

Abstract
A polymorphism in the promoter of human NRAMP1 encodes a Z-DNA forming dinucleotide repeat with four alleles: (1) t(gt)5ac(gt)5ac(gt)11g; (2) t(gt)5ac(gt)5 ac(gt)10g; (3) t(gt)5ac(gt)5ac(gt)9g; and (4) t(gt)5ac(gt)9g. Alleles 1 and 4 are rare (gene frequencies ∼0.001); alleles 2 and 3 occur at gene frequencies ∼0.20−0.25 and ∼0.75− 0.80, respectively. Here, luciferase reporter gene constructs are used to show that the four alleles differ in their ability to drive gene expression. In the absence of exogenous stimuli, alleles 1, 2, and 4 are poor promoters; allele 3 drives high expression, indicating that the repeat itself has endogenous enhancer activity. All four alleles show a similar percentage enhancement of reporter gene expression in the presence of interferon-γ, consistent with the multiple interferon-γ response elements both 5′ and 3′ of the Z-DNA forming repeat. However, while the addition of bacterial lipopolysaccharide (LPS) has no effect on alleles 1 and 4, it causes significant reduction in expression driven by allele 2 and enhances expression driven by allele 3, suggesting that the juxtaposition of LPS related response elements (NFκB, AP-1, NF-IL6) may be differentially affected by the two commonly occurring alleles. These results are consistent with the hypothesis that chronic hyperactivation of macrophages associated with allele 3 is functionally linked to autoimmune disease susceptibility, while the poor level of NRAMP1 expression promoted by allele 2 contributes to infectious disease susceptibility. Conversely, allele 3 protects against infectious disease and allele 2 against autoimmune disease. Hence, alleles that are detrimental in relation to autoimmune disease susceptibility may be maintained in the population because they improve survival to reproductive age following infectious disease challenge.
- human NRAMP1 gene
- autoimmune disease susceptibility
Statistics from Altmetric.com
The murine macrophage resistance gene Nramp1 regulates priming/activation of macrophages for enhanced microbicidal/tumoricidal activity. The pleiotropic effects of Nramp1 on macrophage function,1 which include enhanced chemokine KC, tumour necrosis factor α, interleukin 1β, inducible nitric oxide synthase, and MHC class II expression, are all of potential importance in the induction/maintenance of autoimmune diseases such as rheumatoid arthritis, but are also crucially important in resistance to intramacrophage pathogens such as tuberculosis. We previously showed2 that the promoter region of NRAMP1 possesses a polymorphism within a possible enhancer element containing a Z-DNA forming dinucleotide repeat. Four alleles were identified: (1) t(gt)5ac(gt)5ac(gt)11g; (2) t(gt)5ac(gt)5ac(gt)10g; (3) t(gt)5ac(gt)5ac(gt)9g; and (4) t(gt)5ac(gt)9g (this allele was incorrectly reported as t(gt)5 ac(gt)5ac(gt)4g in the original report2). Alleles 1 and 4 are rare (gene frequencies ∼0.001) in all populations. Allele 2, which typically occurs at a gene frequency of ∼0.20−0.25 in white,3 4Brazilian,5 or African6 populations, is associated with infectious (tuberculosis) disease susceptibility.6 7 Allele 3, which occurs at a gene frequency of ∼0.75−0.80, is associated with autoimmune disease susceptibility: rheumatoid arthritis,3 juvenile rheumatoid arthritis,4 and diabetes in patients with a first or second degree relative with rheumatoid arthritis.8Crohn’s disease has also recently been shown to be associated with polymorphisms closely linked to the NRAMP1 gene,9 but the NRAMP1 promoter region polymorphism was not examined.
The aim of the present study was to use luciferase reporter gene constructs to determine whether the naturally occurring promoter region alleles drive different levels of NRAMP1 expression, and to relate this functionally to the opposing allelic associations observed between NRAMP1 and susceptibility to infectious versus autoimmune diseases.
Materials and methods
CELL CULTURE
Undifferentiated human myeloid cell line U937 was maintained in RPMI-1640 supplemented with 10% fetal calf serum (Gibco BRL) and 2 mmol/l L-glutamine (Sigma).
SEQUENCING OF 5′ FLANKING REGION OF NRAMP1
An Alu based PCR was used to obtain a product which could be used to extend the promoter region sequence 5′ of the previously reported2 yeast artificial chromosome (YAC) derived sequence. The Alu PCR was performed on DNA from this YAC using a non-specific Alu primer paired with an internal primer (5′CCGGTTTTGTGTCTGGGAT3′) to yield a product of 2337 bp. This product was TA cloned (Invitrogen BV, The Netherlands) and primer walking used to extend the sequence.
CONSTRUCTION OF PLASMIDS
A series of luciferase reporter gene constructs were generated in the pGL2-Basic vector (Promega Corp, Southhampton, UK) for the four alleles associated with the GT repeat polymorphism present in the 5′ flanking region of human NRAMP1. The constructs for alleles 1, 2, 3, and 4 incorporated 587, 585, 583, or 571 bp 5′ of the ATG initiation codon, respectively. Each allele was cloned into the pGL2-Basic vector 5′ of the initiation codon for the reporter gene luciferase. The constructs were designated Luc1 to Luc4. Each allele was PCR amplified using high fidelity Vent polymerase (New England Biolabs, Hitchin, UK) from YAC (allele 3) or genomic (alleles 1, 2, 4) DNA using primers (5′AGTGCTAGCCTGCAGTGCCTTCCTC TG3′, 5′TGAAGATCTGCCGACTTCAGGTACTCT3′; positions of the specific sequence indicated in fig 1) designed to incorporate NheI andBglII restriction sites. PCR products were initially cloned into the TA cloning vector pCRII (Invitrogen BV, The Netherlands) and their sequence verified before excision withNheI and BglII and cloning into the pGL2-Basic vector.
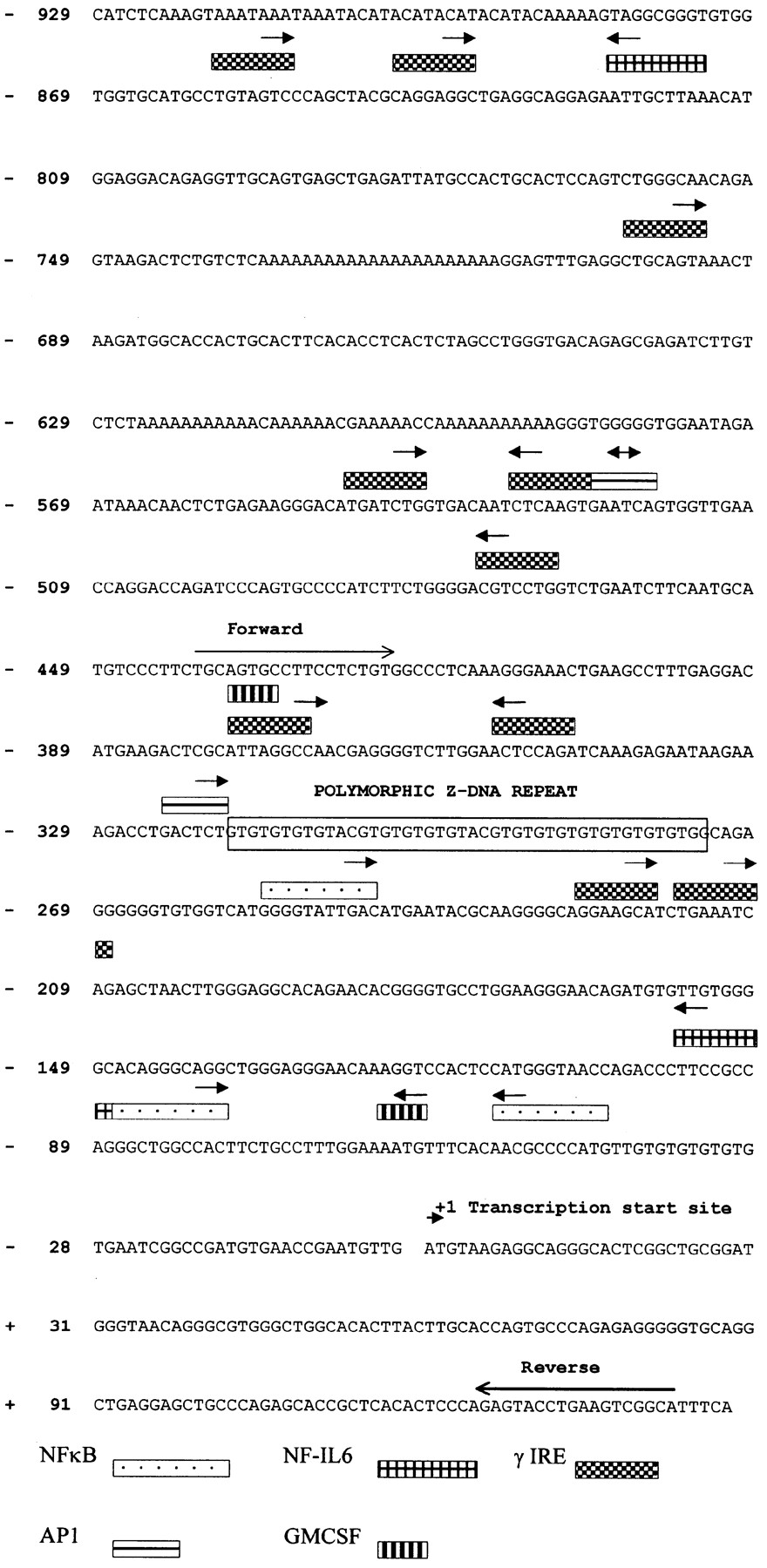
Sequence for the promoter region of human NRAMP1, showing 948 bp of sequence 5′ of the transcription start site, extending the original2 440 bp of sequence by 508 bp and identifying seven γ-IREs, one NF-IL6, and one AP1 site additional to those previously reported. The sequence is also annotated with two GMCSF elements and one NF-IL6 element not included in the original report. The positions of PCR primers used to amplify segments used for the reporter gene constructs are also indicated.
TRANSIENT TRANSFECTIONS AND LUCIFERASE ASSAYS
Log phase U937 cells grown in fully supplemented medium were transfected with the reporter constructs. Cells were resuspended at 2 × 107/ml and 400 μl of the suspension cotransfected by electroporation (300 volts, 900 uF, and resistance α; EasyjecT plus electroporator, Flowgen, Lichfield UK) with 10 μg Luc NRAMP1 construct and 2 μg of a control β-galactosidase construct (kindly provided by Dr A Green, Cambridge, UK). Transfected cells were resuspended in 5 ml of fully supplemented medium and incubated (37°C, 5% CO2) for six hours with/without addition of the exogenous stimuli interferon-γ (100 units/ml) alone or interferon-γ (100 units/ml) plus bacterial lipopolysaccharide (LPS, 10 ng/ml). Lysates were prepared for assay of luciferase and β-galactosidase activities. The cells were harvested, lysed in a buffer containing 25 mmol/l glycyl-glycine, pH 7.8, 15 mmol/l MgSO4, 4 mmol/l ethyleneglycol bis-tetracetic acid (EGTA), 1% vol/vol Triton X100 and 1 mmol/l DTT for 10 minutes at room temperature. Extracts were assayed for luciferase activity in a buffer containing 25 mmol/l glycyl-glycine, 15 mmol/l MgSO4, and 5 mmol/l ATP. Then 100 μl of cell lysate was added to 300 μl of assay buffer and read in an Autolumat LB 953 luminometer (Berthold, Stevenage, UK). The β-galactosidase assay was performed on 10 μl of cell lysate in 150 μl of buffer containing 1 mmol/l MgCl2, 50 mmol/l β-ME, 120 μg of ONPG (o-nitrophenyl-β-δ galactopyranoside), and 7.4 mmol/l sodium phosphate buffer, pH 7.3, and read (absorbance 414 nm; Multiskan, Labsystems, Basingstoke, UK) following a 30 minute incubation at 37°C. To take account of variations in transfection efficiency for different reporter gene constructs, relative light units (RLU) of luciferase activity were normalised against the β-galactosidase reading for each construct within an experiment. Absolute values for normalised relative light units were comparable within but not between experiments. Duplicate transfections were performed for each construct and set of conditions within each experiment. In all cases, duplicate readings fell within the mean ± 20%. To compare data over multiple experiments, normalised RLU values for Luc NRAMP1 constructs were expressed relative to the pGL2-Basic vector control included in every experiment. The two tailed, unpaired Student’st test was used to test for significant differences between constructs and treatments.
Results
EXTENSION OF THE 5′ FLANKING REGION SEQUENCE OF NRAMP1
The primary aim of the present study was to use reporter gene constructs to determine whether polymorphisms previously identified in the NRAMP1 promoter region2 are functionally important in differential regulation of gene expression. In order to create constructs with adequate flanking sequence 5′ of this polymorphic region, additional 5′ sequence data was required. Fig 1shows 948 bp of promoter region sequence 5′ of the transcription start site, extending the original 440 bp by 508 bp and identifying seven interferon-γ response elements (γ-IRE), one NF-IL6 site, and one AP1 site additional to those previously reported.2 The original 440 bp of sequence is also annotated (fig 1) with two GMCSF elements and one NF-IL6 element not included in the original report. All of these elements are consistent with studies in the mouse showing that expression of Nramp1 is regulated by GMCSF,10interferon-γ,10-12 and LPS.11
DEMONSTRATION THAT THE FOUR NRAMP1 PROMOTER REGION ALLELES DRIVE DIFFERENT LEVELS OF REPORTER GENE EXPRESSION
Initial experiments were designed to determine whether the four naturally occurring promoter region alleles differed in their ability to drive reporter gene expression without the addition of an exogenous stimulus. The mean (SD) for quadruplicate data points obtained over two independent transfection experiments in which the four Luc NRAMP1 constructs were tested are shown in fig 2. Allele 3 was observed to drive up to 10-fold higher levels of luciferase reporter gene expression than each of alleles 1, 2, or 4. Over the two experiments, differences between allele 3 and each of alleles 1, 2, and 4 were statistically significant (two tailed, unpairedt test) at p=0.008, p=0.001, and p=0.018, respectively. Allele 1 also differed significantly from alleles 2 (p=0.004) and 4 (p=0.022). The pGL2-Basic vector lacks any eukaryotic, promoter, or enhancer elements. Hence, luciferase activity measured following transient transfections with the reporter gene constructs represents the action of the inserted NRAMP1 promoter region. Since the only difference between the four constructs is in the repeat structure, the differential expression observed without the addition of an exogenous stimulus indicates that the Z-DNA forming repeat is itself acting as an enhancer element.

Levels of luciferase reporter gene activity driven by the four naturally occurring polymorphic alleles for the human NRAMP1 gene promoter. The mean (SD) RLU for quadruplicate data points obtained over two independent experiments are graphed for Luc NRAMP1 constructs relative to the pGL2-Basic vector only control.
EFFECT OF EXOGENOUS STIMULI ON REPORTER GENE EXPRESSION DRIVEN BY THE FOUR POLYMORPHIC NRAMP1 ALLELES
Addition of interferon-γ as an exogenous stimulus causes a one- to threefold enhancement of luciferase reporter gene expression for all four constructs (fig 3). The mean (SD) for quadruplicate data points obtained over two independent transfection experiments in which the four Luc NRAMP1 constructs were tested with/without addition of interferon-γ are shown in fig 3. As before (fig 2), differences in baseline activity between allele 3 and each of the other three alleles were statistically signifnicant (two tailed, unpairedt test) at p<0.001.

Levels of luciferase reporter gene activity driven by the four naturally occurring polymorphic alleles for the human NRAMP1 gene promoter with/without addition of the exogenous stimulus interferon-γ (IFN; 100 U/ml). The mean (SD) RLU for quadruplicate data points obtained over two independent experiments are graphed for Luc NRAMP1 constructs relative to the pGL2-Basic vector only control.
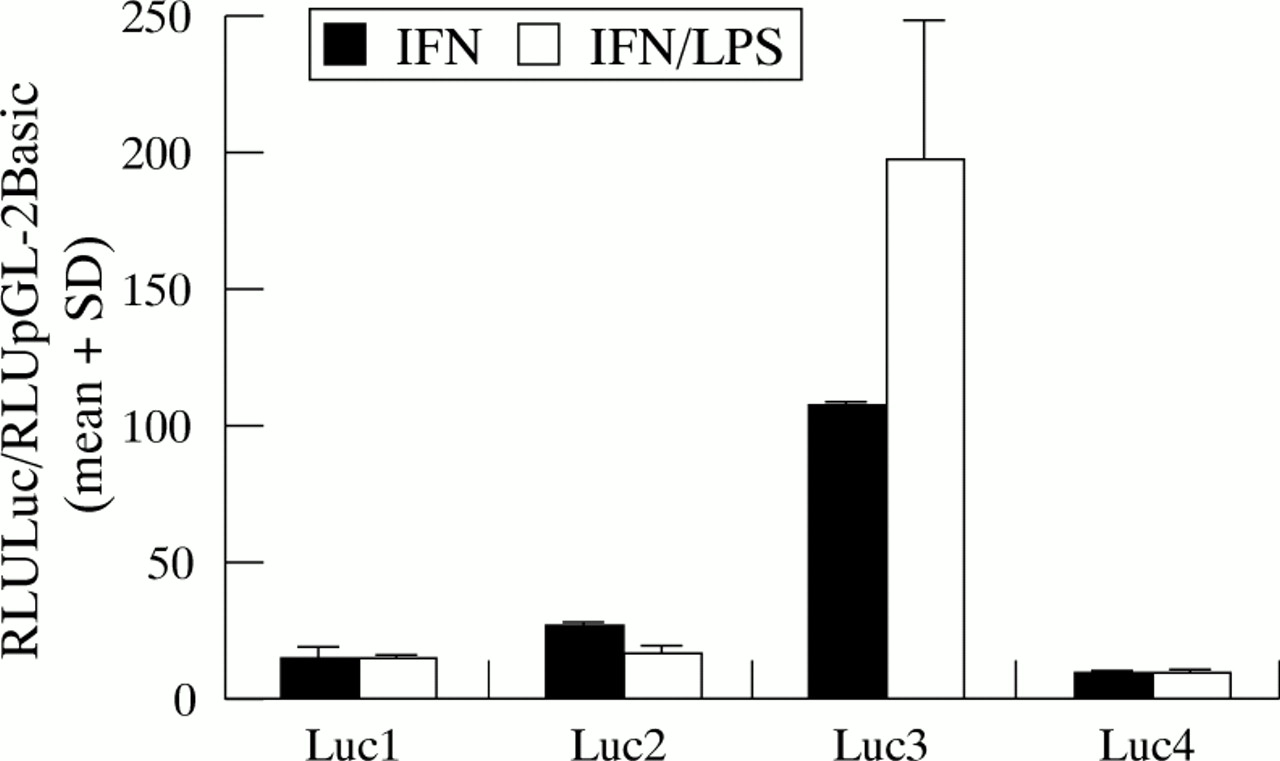
Increases in reporter gene activity with addition of interferon-γ were statistically significant at p=0.0006, p=0.0005, p=0.0004, and p=0.0007 for alleles 1, 2, 3, and 4, respectively, indicating that the multiple γ-IREs both 5′ and 3′ of the Z-DNA forming polymorphic repeat are generally important in upregulating gene expression across all four alleles. On the background of this interferon-γ enhanced response, addition of LPS as a second exogenous signal results in a further ∼twofold enhancement of allele 3 driven reporter gene expression, but causes reduction in the allele 2 driven response (fig4). The mean (SD) for quadruplicate data points obtained over two independent transfection experiments in which the four Luc NRAMP1 constructs were tested with interferon-γ alone or with interferon-γ plus LPS are shown in fig 4. Over the two experiments, there was a significant increase (p=0.013) in reporter gene activity driven by allele 3 with the addition of LPS, and a significant decrease (p=0.0006) in reporter gene activity driven by allele 2 in the presence of LPS. Allele 1 and allele 4 driven responses remain unchanged by the further addition of LPS (fig 4). In a further experiment using only Luc 2 and Luc 3 constructs, a significant increase (p<0.001) in reporter gene activity was again observed with addition of LPS for Luc 3, and a significant decrease (p<0.001) for allele 2. These results suggest that the juxtaposition of LPS related response elements (NFκB, AP-1, NF-IL6) may be differentially affected by the two commonly occurring NRAMP1 promoter region alleles.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Influence of LPS on levels of luciferase reporter gene activity driven by the four polymorphic alleles for the human NRAMP1. The influence of LPS (10 ng/ml) is superimposed on the background of the interferon-γ (IFN; 100 U/ml) enhanced luciferase expression observed across all four alleles (fig 3). The mean (SD) RLU for quadruplicate data points obtained over two independent experiments are graphed for Luc NRAMP1 constructs relative to the pGL-2Basic vector only control.
Discussion
Nramp1 has multiple pleiotropic effects on macrophage activation pathways.1 The gene encodes a polytopic integral membrane protein which is localised to late endosomal and lysosomal compartments13-15 and functions as a metal ion transporter regulating,16 and regulated by,13cellular iron levels. In the mouse, a point mutation in the coding region of the gene results in a single non-conservative amino acid substitution from gly to asp at position 169 in putative17membrane spanning domain number 4, the effect of which is to create a functional null equivalent in infectious disease susceptibility phenotype to genetically engineered gene disrupted mice.18The only potentially detrimental polymorphism identified in the coding region of human NRAMP1, an amino acid triplet repeat in exon 2 encoding the putative SH3 binding domain,19 occurs at low frequency (<0.001; J M Blackwell, unpublished observation) and has never been observed in the homozygous condition. Presumably the NRAMP1 protein in humans is essential to macrophage function, such that detrimental coding region mutations will be rare. On the other hand, polymorphisms which regulate expression of a functional protein may provide a more subtle way in which natural selection influences the role of this important macrophage activation gene in determining disease susceptibility. Here we have shown that the previously identified Z-DNA forming polymorphic repeat in the promoter region of the human NRAMP1 gene does act as a functional polymorphism influencing gene expression, with the commonly occurring alleles 2 and 3 driving dramatically different levels of reporter gene expression.
Several studies now support the hypothesis that this functional repeat polymorphism in the promoter region of human NRAMP1 may contribute in a direct way to the differing allelic associations observed with infectious versus autoimmune disease susceptibilities. In rheumatoid arthritis,3 juvenile rheumatoid arthritis,4and in diabetes patients with a first or second degree relative with rheumatoid arthritis,8 family based transmission disequilibrium testing or case-control analyses have all shown an allelic association between allele 3 of the NRAMP1 promoter region polymorphism and disease susceptibility, while allele 2 is significantly protective. Conversely, our preliminary observation of an association between allele 2 and susceptibility to tuberculosis in 72 genetically independent age and sex matched Brazilians7has now been confirmed in a study of 400 cases and controls from The Gambia in West Africa.6 In the Gambian study, allelic association between the 5′ promoter region of the gene and disease susceptibility was independent of association with polymorphisms in the 3′ region of the gene, indicating that multiple NRAMP1 polymorphisms influence tuberculosis in this population. This could indicate that the repeat polymorphism in the 3′ UTR of the gene20 21 also regulates gene expression. Overall, the data now available on autoimmune and infectious disease associations are consistent with the hypothesis3 that chronic hyperactivation of macrophages associated with high expression promoted by allele 3 may be functionally linked to autoimmune disease susceptibility, while the poor level of NRAMP1 expression promoted by allele 2 leads to infectious disease susceptibility. Together with the functional data presented here, these opposing allelic associations between NRAMP1 promoter alleles and autoimmune versus infectious disease susceptibility suggest that alleles that are detrimental in relation to autoimmune disease susceptibility may be maintained in the population because they improve survival to reproductive age following infectious disease challenge.
Acknowledgments
This work was supported by The Wellcome Trust.