Article Text

Abstract
Disease associated balanced chromosome rearrangements (DBCRs) have been instrumental in the isolation of many disease genes. To facilitate the molecular cytogenetic characterisation of DBCRs, we have generated a set of >1200 non-chimeric, cytogenetically and genetically anchored CEPH YACs, on average one per 3 cM, spaced over the entire human genome. By fluorescence in situ hybridisation (FISH), we have performed a systematic search for YACs spanning translocation breakpoints. Patients with DBCRs and either syndromic or non-syndromic mental retardation (MR) were ascertained through the Mendelian Cytogenetics Network (MCN), a collaborative effort of, at present, 270 cytogenetic laboratories throughout the world. In this pilot study, we have characterised 10 different MR associated chromosome regions delineating candidate regions for MR. Five of these regions are narrowed to breakpoint spanning YACs, three of which are located on chromosomes 13q21, 13q22, and 13q32, respectively, one on chromosome 4p14, and one on 6q25. In two out of six DBCRs, we found cytogenetically cryptic deletions of 3-5 Mb on one or both translocation chromosomes. Thus, cryptic deletions may be an important cause of disease in seemingly balanced chromosome rearrangements that are associated with a disease phenotype. Our region specific FISH probes, which are available to MCN members, can be a powerful tool in clinical cytogenetics and positional cloning.
- chromosomal translocation
- fluorescence in situ hybridisation (FISH)
- Mendelian Cytogenetics Network (MCN)
- mental retardation (MR)
Statistics from Altmetric.com
- chromosomal translocation
- fluorescence in situ hybridisation (FISH)
- Mendelian Cytogenetics Network (MCN)
- mental retardation (MR)
The incidence of de novo balanced chromosome rearrangements among newborns is approximately 1:2000.1 The risk of congenital abnormalities in newborns with such rearrangements is estimated to be 6.1%, compared to a 2-3% risk in random newborn populations.1 2 At least in part, the increased frequency of abnormalities in cases with de novo balanced chromosome rearrangements can be ascribed to disruption or inactivation of specific genes. Molecular characterisation of DBCRs has already led to the identification of several disease genes,3 4 showing the general usefulness of this approach. So far, a systematic effort for the central registration of DBCRs through the Mendelian Cytogenetics Network (MCN, coordinator Niels Tommerup, Denmark) has compiled information on close to 1000 DBCRs provided by approximately 270 participating laboratories. The MCN focuses on the identification and subsequent characterisation of “matching” cases with coincident chromosome breakpoints and similar clinical features.
Roughly one third of all human genes are expressed in the brain5 and many of them are thought to play important roles in brain development and function. Consistent with this, nearly half of the DBCRs are associated with severe or moderate MR. Cryptic deletions and translocations in the gene rich telomeric and subtelomeric regions seem to be an important cause of unexplained MR,6 but other as yet unidentified chromosome regions and genes must also be involved. Therefore, fine mapping and cloning of cytogenetically balanced chromosome breakpoints in patients with MR is a promising strategy for the isolation of new MR genes.
As a tool for systematically studying such DCBRs, we have generated a large set of cytogenetically and genetically anchored YACs, which are more or less evenly spaced and cover approximately one third of the human genome. We have studied six unrelated DCBRs that are associated with syndromic or non-syndromic MR. The chromosome bands containing the specific breakpoints are involved in at least three independent MR associated balanced chromosome rearrangements, which are recorded in the MCN database. In a short time, we have identified five YACs spanning MR associated translocation breakpoints. Moreover, we have shown that a significant number of seemingly balanced de novo translocations show cryptic deletions which almost certainly account for the observed clinical phenotype.
Methods
CASE REPORTS
Blood or cells from the following patients and their parents were provided by the MCN. In all cases, routine cytogenetic banding showed de novo, apparently balanced, reciprocal chromosome translocations.
Case 1: 46,XY,t(6;13)(q25;q22)
This boy was born at term in 1992 to a 29 year old mother and a 32 year old father. He was the fourth child of healthy, non-consanguineous parents. There was no history of spontaneous abortion. Birth weight of the infant was 3000 g (25th centile), length 50 cm (25th centile), and head circumference 35 cm (50th centile). The following clinical findings were noted: facial dysmorphism including hypertelorism, slight antimongoloid position of the eyes, low set ears, thick lips, and long philtrum. Later examination showed bilateral cryptorchidism and an umbilical hernia. At 16 months his height was 72 cm (<3rd centile) and his weight was 7350 g (<3rd centile). He sat at 12 months and walked at 3 years. Clinical re-examination at 4 years of age showed hypotonia and MR. He walked with difficulty and his language ability was poor.
Case 2: 46,XX,t(3;13)(q21;q22.2)
The patient was born in 1984 with a birth weight of 3400 g (50th centile) to a 31 year old mother and a 30 year old father. The family’s first child had been a girl with MR and a 46,XX,8p+ karyotype. Subsequent pregnancies resulted in two spontaneous abortions, followed by the birth of the patient and a healthy younger brother. The parents observed developmental delay in the infant, with sitting at 8 months and walking at 30 months. Clinical analysis of the patient at the age of 5 years showed delayed psychomotor development and hypotonia. Language skills were very poor. Her weight was 18.5 kg (50th centile), height 109.5 cm (50th-75th centile), and head circumference 52 cm (75th centile). Kyphoscoliosis of the spine was noted. She had frequent bronchitis and diarrhoea. Clinical re-examination at the age of 12 showed moderate MR and poor language skills.
Case 3: 46,X,t(X;4)(p21;p14),inv(2)(p15q11.1),t(4;13) (q21;q22)
This complex karyotype is associated with profound MR in the patient. The patient was born in 1957 and noted to have simian creases. She is physically normal except that she has a low frontal hair line and abnormally short hands.
Case 4: 46,XY,t(4;13)(p14;q32)
This boy was born in 1978 with a birth weight of 3025 g (25th centile) to a 30 year old mother and a 36 year old father. In the neonatal period, septic shock, acute renal insufficiency, and anaemia occurred. Clinical examination at 3 months showed a weight of 4500 g (10th centile) and a height of 60 cm (25th centile). His head circumference was 38 cm (3rd centile). Cardiopulmonary examination and urography were normal. EEG was within normal limits, but low voltage with diffuse cerebral dysfunction was noted. By the age of 2 years, he showed signs of MR (Gessell test: motor 53%, adaptive 53%, speech 42%, social 53%). He walked at 22 months. At the age of 4 years he had generalised seizures and he was very nervous and displayed stereotypic movements and strabismus. At 18 years his weight was 90 kg (>97th centile) and his height was 182 cm (75th centile). He could eat and dress alone but at a special school he did not make any progress.
Case 5: 46,XY,t(6;13)(p21.2;q33.3)
The patient was born in 1986 to a 32 year old mother with a birth weight of 3500 g (50th centile). Pregnancy and delivery were normal. The father’s age is not known. The two older brothers are healthy. By the age of 2 months, the infant showed delayed psychomotor development. He had large, low set ears, microcephaly, and strabismus convergens. From the age of 6 years he suffered from epilepsy. At this time he was diagnosed with moderate MR. CT scan was normal.
Case 6: 46,XY,t(7;8;11;13)(q21.1;q21.3;p14.3;q21.2)
This patient, who has classical Moebius syndrome (MBS, MIM 157900), carries a complex reciprocal translocation involving four different chromosomes. He was born in 1958 to a 35 year old father and a 32 year old mother. The family already had two healthy children. Because of a paresis of the facial muscles as a newborn, he had feeding problems owing to inefficient sucking and swallowing. A more detailed examination at the age of 17 showed moderate to severe MR and deficient language development. At this time, his height was only 156 cm and his weight 39.5 kg (<3rd centile). Other clinical features included ptosis, blepharophimosis, atrophy of the optic nerve, and severe amblyopia. His mandible and facial muscles were hypoplastic, leading to difficulties in opening his mouth. His metacarpals and metatarsals were short, limiting the mobility of his hands and feet. Pectoral muscles were hypoplastic.
DNA PROBES
YAC clones of interest were selected from the CEPH mega-YAC library and obtained through the Reference Library Database system (Berlin, Germany). Details on genetic markers of individual YACs were obtained through public databases (CEPH/Généthon,http://www.cephb.fr/bio/ceph-genethonb-map. html; Whitehead Institute,http://www.genome. wi.mit.edu). The human inserts were isolated by pulsed field gel electrophoresis (PFGE) and amplified by degenerate oligonucleotide primed polymerase chain reaction (DOP-PCR) according to the protocol of Telenius et al 7with minor modifications.
CHROMOSOME PREPARATION
Metaphase chromosomes were prepared from human peripheral blood lymphocytes and from Epstein-Barr virus transformed lymphoblastoid cell lines and 0.1 μg/ml of colcemid (Gibco) was added to the culture medium one hour before cell harvest to arrest cells in metaphase. Cell pellets were resuspended in a hypotonic solution consisting of 50 mmol/l KCl. After 20 minutes of hypotonic treatment, cells were fixed overnight with 3:1 methanol:acetic acid. Slides were prepared using the conventional drop-splash technique.
FLUORESCENCE IN SITU HYBRIDISATION
Standard FISH protocols were followed.8 DOP-PCR products of PFGE purified YAC inserts were labelled with either biotin-16-dUTP or digoxigenin-11-dUTP (Boehringer Mannheim) by standard nick translation. The slides were treated with 100 μg/ml RNase A in 2 × SSC, pH 7.0, at 37°C for one hour and with 0.01% pepsin in 10 mmol/l HCl at 37°C for 10 minutes, then rinsed three times in 2 × SSC, and dehydrated in an ethanol series (70%, 80%, 100%). Slides were denatured at 80°C in 70% formamide, 2 × SSC, pH 7.0, and again dehydrated in an alcohol series. The hybridisation mixture was composed of 50% formamide in 2 × SSC, 10% dextran sulphate, 500 μg/ml salmon sperm DNA, 100 μg/ml cot-1 DNA (per YAC), and 10 ng/μl biotinylated or digoxigenated DNA of each YAC. Preannealing of repetitive sequences with cot-1 DNA (Gibco BRL) was carried out for 20 minutes at 37°C before application to the denatured metaphase spread. After five minutes denaturation at 70°C, 30 μl of the hybridisation mixture was applied to each slide and sealed under a coverslip. Hybridisation was done overnight in a moist chamber at 37°C. The slides were then washed twice for five minutes in 50% formamide, 2 × SSC at 42°C and twice for five minutes in 0.1 × SSC, pH 7.0, at 60°C. After washing, the probes were detected with fluorescein isothiocyanate (FITC) conjugated avidin (Vector Laboratories) and Cy3 conjugated antidigoxigenin antibody (Dianova). Chromosomes and cell nuclei were counterstained with 1 μg/ml 4′,6-diamidino-2-phenylindole (DAPI) in 2 × SSC for three minutes. The slides were mounted in 90% glycerol, 0.1 mol/l Tris-HCl, pH 8.0, and 2.3% 1,4-diazobicyclo-2,2,2-octane.
DIGITAL IMAGING MICROSCOPY
Images were taken with a Zeiss epifluorescence microscope equipped with a thermoelectronically cooled charge coupled device camera (Photometrics CH250), which was controlled by an Apple Macintosh computer. Oncor imaging software was used to capture grey scale images and to superimpose the images creating a colour image. Oncor imaging software was also used to convert the DAPI image into a G banded metaphase for identification of the chromosomes.
Results
As a prerequisite for the systematic molecular cytogenetic characterisation of DBCRs, we have selected a standard set of cytogenetically and genetically anchored CEPH YACs. To generate complex FISH probes on a larger scale, PFGE isolated human YAC inserts were amplified by DOP-PCR. Because of their large insert size (of at least several hundred kilobases), CEPH YACs yield high intensity FISH signals on both metaphase spreads and interphase nuclei. For all YACs in our probe panel, physical mapping information and sequence tagged site (STS) markers were available. In view of its higher resolution, probe ordering was based on relative positions of YAC markers on the genetic map.9 FISH mapping of genetically anchored YACs on metaphase chromosomes allows rapid integration of genetic and cytogenetic maps.10-12 Each individual YAC clone was examined for chimerism by FISH and linked with STS markers to a specific chromosome region. This allows one to select FISH probes for any genomic region of interest either on the basis of available genetic information or chromosomal location. Details about the growing set of probes and their availability are provided at the Molecular Cytogenetics and Positional Cloning Center internet site (http://www.mpimg-berlin-dahlem.mpg.de). Additional information on STS markers and YAC contigs can be retrieved from the Whitehead Institute database (http://www.genome.wi. mit.edu).
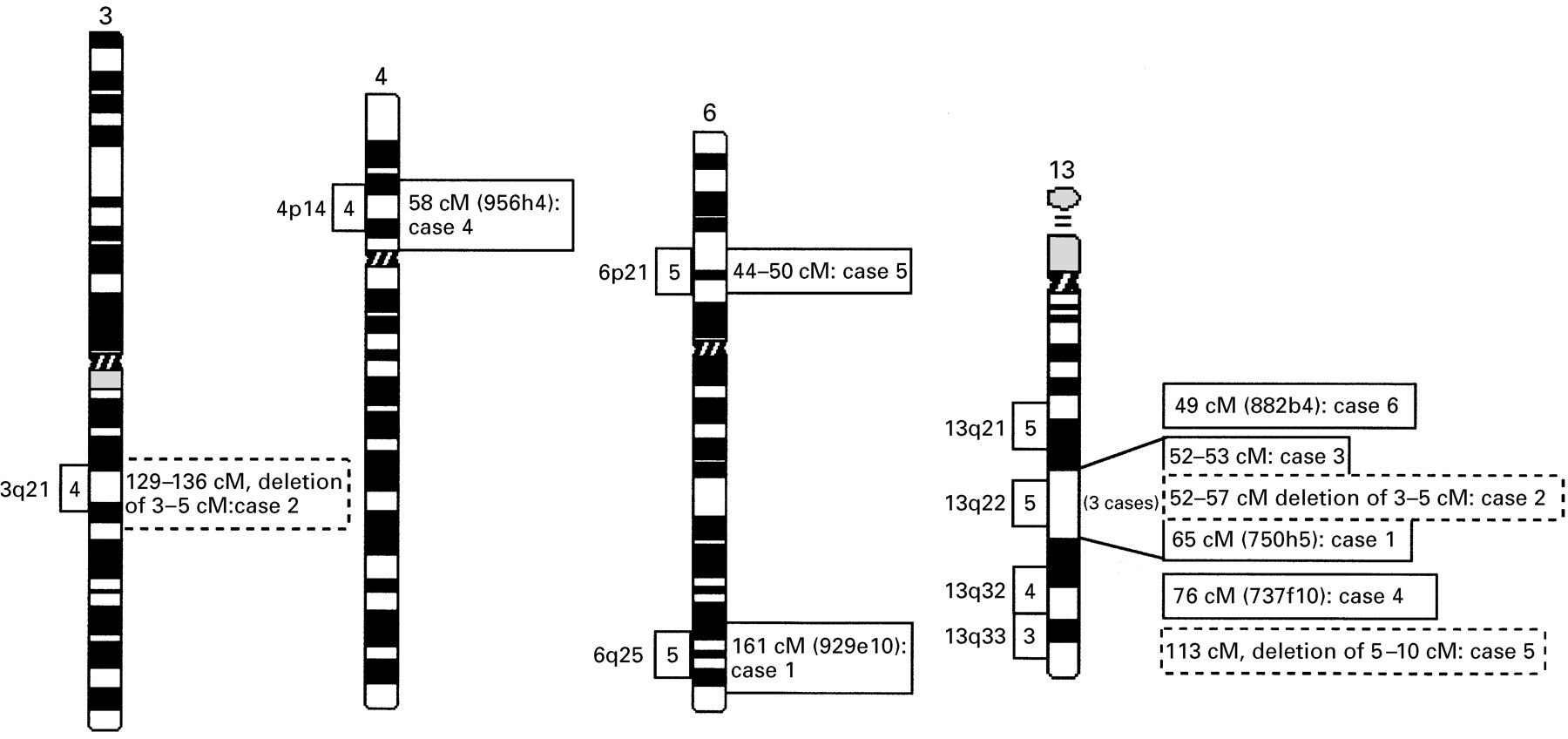
Screening of the MCN database showed that more than 500 DBCRs associated with MR are clustered in different chromosome bands or regions (N Tommerup, personal communication). In this pilot study, a total of four different breakpoint regions on chromosome 13 and four regions on other chromosomes were investigated. These translocation cases were chosen because the respective breakpoint regions are involved in at least three independent MR associated balanced chromosome rearrangements (fig 1). It is probable, therefore, that these regions harbour genes which are important for brain development and function.
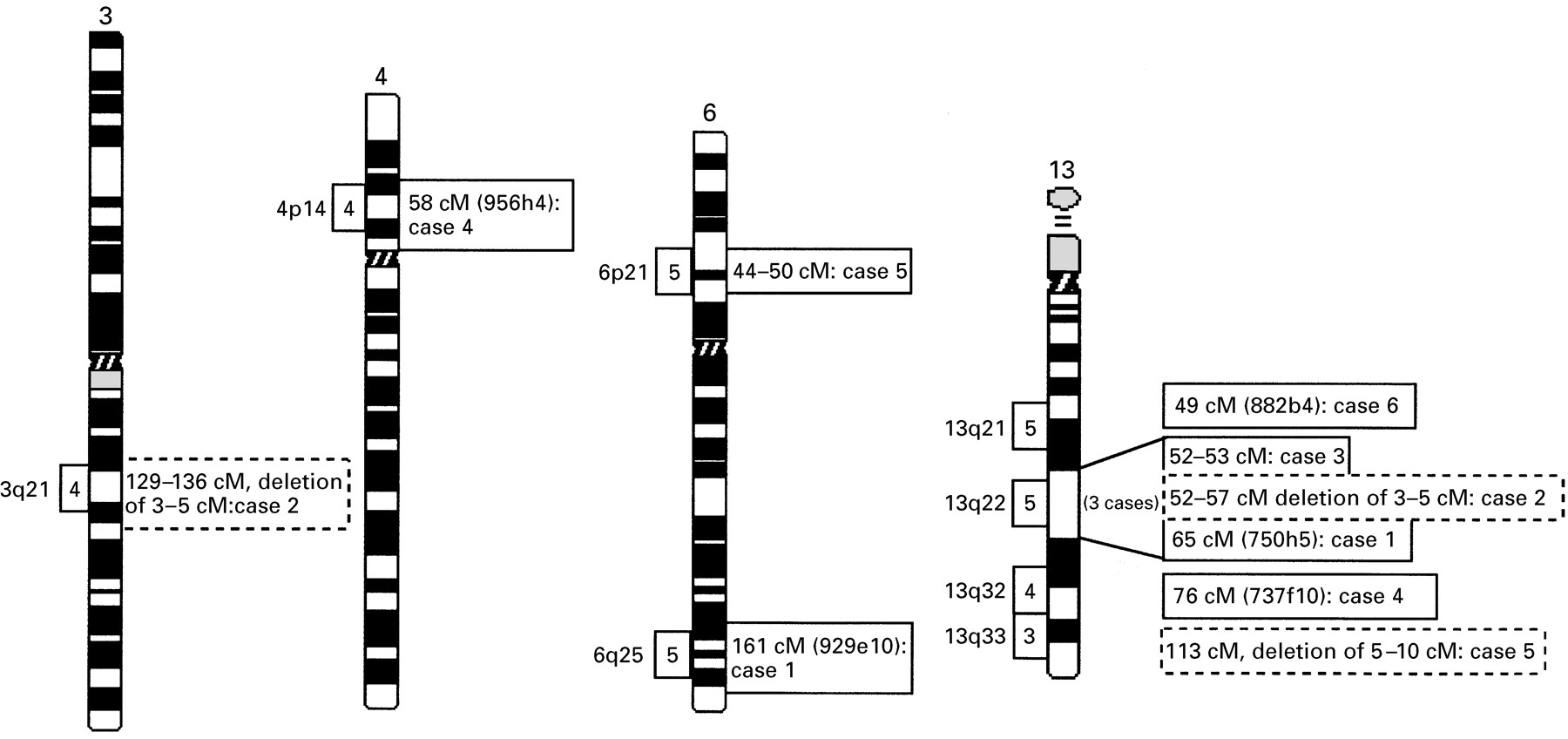
Location of MR associated translocation breakpoints on chromosome 3, 4, 6, and 13. Framed numbers (3, 4, 5) on the left side of a chromosome region indicate the number of MR associated breakpoints within this specific band that are registered in the MCN database. The corresponding figures on the right side of each region indicate the genetic localisation of the different breakpoints studied here. Genetically anchored YACs spanning the relevant breakpoints are shown in brackets. Breakpoints associated with deletions are indicated by dashed frames. The distances in cM are given on the right.
MR associated balanced rearrangements are clustered on the long arm of chromosome 13 (fig 1). Cytogenetically, three breakpoints of different translocation cases (our cases 1, 2, and 3) had all been assigned to the same chromosome band, 13q22. However, when YAC mapping was applied, the breakpoints proved to be located in different subregions (figs 1and 2). Several region specific YAC probes were used to map the 13q22 breakpoint of case 1 with a t(6;13). The breakpoint spanning YAC 750h5 (D13S170, 65 cM) clearly hybridised to both the der(13) and the der(6) (fig 2A). Additionally, a subset of seven different YACs were FISH mapped and showed regular signals on one of the derivative chromosomes and the normal chromosome 13 (fig 2B). The 6q breakpoint of case 1 was defined by the breakpoint spanning YAC 929e10 (D6S1633, 161cM). Additional YACs mapping either proximal or distal to this breakpoint indicate that the chromosome 6 rearrangement is balanced as well (fig2B).

FISH mapping of YACs on the chromosomes of translocation cases 1, 2, and 3 (A, C, and E). Locations of YACs for each translocation case are noted as proximal, distal, or spanning. Asterisks denote chimeric YACs (B, D, and F). (A) Case 1: t(6;13)(q25;q22): YAC 921f2 (green) is proximal to the 13q22 breakpoint with signals seen on the normal chromosome 13 and the der(13). The breakpoint spanning YAC 750h5 (red) gives signals on both rearranged chromosomes, the der(13) and the der(6). (C) Case 2: t(3;13)(q21;q22.2): YAC 908b1 (green) gives signals on the normal chromosome 13 and the der(13). For YAC 758b1 a red signal is seen only on the normal chromosome 13, but not on the rearranged chromosomes, der(13) and der(3), indicating a deletion in the breakpoint region. (E) Case 3: t(4;13)(q21;q22)complex: YAC 908b1 (green) and YAC 758b1 (red) map very close to each other on the normal chromosome 13. On the rearranged chromosomes the green signal is retained proximally on the der(13), while the red signal is seen distally on the der(4).
Unexpectedly, analysis of case 2 with an apparently balanced t(3;13) showed an interstitial cryptic deletion at the 13q22 breakpoint region (fig 2C, D). The proximal deletion breakpoint was mapped between D13S1317 (52 cM) and D13S275 (53 cM), the same genetic interval that contains the breakpoint of case 3 (fig 1, fig 2D). Three YACs, 758b1 (D13S275, 53 cM), 757a8 (D13S279, 54 cM), and 909g3 (D13S152, 56.6 cM), which yielded hybridisation signals on the normal chromosome 13, did not produce signals on the derivative chromosome (fig 2C). YAC 949e1 (D13S166, 57 cM) showed regular signals on both the normal chromosome 13 and the der(3), thus defining the distal boundary of the 13q22 deletion (fig 2D). Since all YAC probes are integrated in the Whitehead contig WC13.3, it could be determined that the deleted region covers 3-5 Mb between D13S275 (53 cM) and D13S152 (56.6 cM). Upon closer examination, the reciprocal translocation product showed an additional cryptic deletion of 3-5 Mb on chromosome 3q21. Sequences homologous to YACs 965b8 (D3S3665, 132 cM), 770c10 (D3S1278, 132 cM), 960f11 (D3S3529, 134 cM), and 767e12 (D3S3529, 134 cM) (fig 2D), were absent from the 3q21 breakpoint region. The proximal and distal breakpoints of the 3q21 deletion are defined by YACs 960d1 (D3S2496, 129 cM) and 967f11 (D3S1558, 136 cM), which showed regular signals on the translocation chromosomes.
When the cytogenetically matching 13q22 breakpoint of case 3 was probed with different YACs, it was found to map to a different subregion (fig2F). The breakpoint region was refined to a 1 cM interval between YAC 908b1 (D13S1317, 52 cM) and 758b1 (D13S275, 53 cM) (fig 2E, F).
Clustering of DBCRs is also observed on the distal long arm of chromosome 13 (fig 1), which is why the 13q32 breakpoint of a patient with a t(4;13) (case 4) was studied in detail. Several YACs containing STS markers between D13S264 (66 cM) and D13S1267 (84 cM) were hybridised to chromosomes of this patient. As shown in fig 3A, split signals on the der(13) and the der(4) were seen with YAC 737f10 (D13S1241, 77 cM). This 13q32 breakpoint maps approximately 5 cM proximal to the critical region for the so-called 13q deletion syndrome, which involves severe brain and digital malformations.13

{kind=link}
{kind=link}
{kind=link}
FISH mapping of YACs on the chromosomes of translocation cases 4, 5, and 6 (A, C, and E). Locations of YACs for each translocation case are noted as proximal, distal, or spanning. Asterisks denote chimeric YACs (B, D, and F). (A) Case 4: t(4;13)(p14;q32): YAC737f10 (red) is spanning the breakpoint. Signals are present on the normal chromosome 13 and both derivative chromosomes, the der(13) and the der(4). YAC 761g6 (green) is distal to the 13q32 breakpoint with signals on the der(4). (C) Case 5: t(6;13)(p21.2;q33.3): YAC 931g12 is proximal to the 13q33.3 breakpoint, producing green signals on the der(13). For the telomeric YAC 908c3 (red) signals are seen only on the normal chromosome 13, indicating a deletion at the 13q33.3 breakpoint region. (E) Case 6: t(8;13)(q21.3;q21.2)complex: FISH results of the breakpoint spanning YAC 882b4 (green). Signals are present on the normal chromosome 13 and both derivative chromosomes, the der(13) and the der(8).
The reciprocal breakpoint on chromosome 4p14 was characterised with five different YACs, 944c7 (64 cM), 931c2 (36 cM), 796d3 (38 cM), 747d6 (51 cM), and 956h4 (58 cM) (fig 3B). The 4p14 breakpoint is contained in YAC 956h4, which is genetically anchored to D4S405 (57 cM) and D4S2919 (58 cM). No cryptic deletions were detected in either breakpoint region.
Case 5 with a t(6;13) showed a breakpoint in the subtelomeric 13q33.3 region and one in 6p21.2. YAC mapping of the breakpoint region 13q33.3 showed a cryptic deletion of sequences homologous to YAC 908c3 (D13S261, 113 cM) (fig 3C, D). This YAC represents the most distal probe of our chromosome 13 panel and is closely juxtaposed to the actual chromosome end.12 YAC 931g12 (D13S173, 96 cM) displayed signals on both the normal chromosome 13 and the der(13) and, therefore, is proximal to the breakpoint (fig 3C). This finding is consistent with the recent observation that a significant percentage of patients with unexplained MR may be accounted for by cytogenetically cryptic deletions in the gene rich (sub)telomeric chromosome regions.6 By FISH there was no evidence for a deletion at the 6p21.2 breakpoint; several region specific YACs (fig 3D) hybridised either proximal or distal to the breakpoint on the derivative chromosomes.
Several region specific YAC probes were used to map the breakpoint on chromosome 13q21.2 of patient 6 who has Moebius syndrome and MR (fig3F). YAC 882b4 located on chromosome 13q14.3-21.1 displayed signals on both the der(13) and the der(8), clearly overlapping the13q21 breakpoint (fig 3E). Because this YAC contains the STS markers D13S1273 (49 cM) and D13S172 (50 cM), the breakpoint of case 6 was placed on the genetic map between 49 cM and 50 cM. All other YAC probes gave regular signals either proximal or distal to the breakpoint in 13q21 (fig 3F), which makes a larger deletion unlikely.
Discussion
In this study, YAC mapping was applied in a systematic and time saving manner to characterise the breakpoints of six apparently balanced translocations that are associated with MR. To streamline the molecular cytogenetic analysis of DBCRs and to facilitate breakpoint cloning, we have generated a set of >1200 non-chimeric, cytogenetically and genetically anchored YAC probes, on average one per 3 cM, placed over the entire human chromosome complement. Cytogenetic laboratories participating in the MCN have filed more than 1000 DBCRs associated with MR or congenital malformations or both. In many cases, FISH with region specific probes will be the method of choice to characterise the respective chromosome breakpoints further. Our YAC panel provides an excellent tool for the large scale study of DBCRs. All the YACs of our probe panel are well characterised with regionally mapped STS markers and expressed sequence tags (ESTs) that have been physically mapped on chromosomes. This information and the fact that many YACs are integrated into larger contig(s)9 should be of help for the molecular characterisation of DBCRs and the isolation of the relevant candidate disease genes. Our region specific FISH probes are available upon request to all MCN members (for details, seehttp://www.mpimg-berlin-dahlem.mpg.de).
Using this unique resource for human molecular cytogenetics, in this study we have focused on 10 different chromosome regions where breakpoints of MR associated balanced chromosome rearrangements are clustered. For five of these regions, breakpoint spanning YACs were identified, three of which are located on chromosome 13q21, 13q22, and 13q32, respectively, one is on chromosome 4p14, and one on 6q25.
The 13q22 breakpoints of three cytogenetically matching MR cases proved to be different at the molecular level, which illustrates that the identification of matching cases on the basis of chromosome banding alone may be misleading. All three patients with 13q22 breakpoints were mentally retarded, but did not exhibit other clearly defined clinical features. Case 2 displayed cryptic deletions of 3-5 Mb in both breakpoint regions. To our knowledge, this is the first description of a de novo rearrangement with cryptic deletions on both reciprocal translocation chromosomes. The proximal breakpoint of the 13q22 deletion (case 2) and the balanced translocation breakpoint of our case 3 both map between YAC 908b1 and 758b1. Therefore, it is still possible that the same gene is affected in both patients. Another cryptic deletion was found in the subtelomeric region 13q33.3 of case 5 with t(6;13)(p21.2;q33.3). These results and data from a larger number of unpublished cases (in preparation) clearly indicate that in a significant proportion (>10%) of cytogenetically balanced translocations, submicroscopic deletions of several megabases account for the associated clinical phenotype. Molecular characterisation of such microdeletions will allow one to define critical regions for MR and other congenital anomalies in the human genome.
It has recently been shown that several percent of patients with unexplained mental retardation have cytogenetically cryptic chromosome rearrangements within the subtelomeric regions,6 and it has been speculated that chromosome ends may be more prone to deletions and translocations because of their increased rate of genetic recombination.6 Indeed, our data suggest that in DBCRs involving the chromosome ends, microdeletions may be relatively common. Hence, apparently balanced translocation breakpoints in (sub)telomeric chromosome bands should be analysed with particular care, using region specific FISH probes.
Cytogenetically visible deletions involving chromosome band 13q32 have been described in certain patients with severe brain and digital malformations.13 The common deletion region in this 13q deletion syndrome encompasses a 1.5 Mb interval between D13S136 and D13S147.13 Furthermore, several other breakpoints of balanced translocations associated with MR were localised to band 13q32 by conventional cytogenetic studies.14 The breakpoint of our t(4;13)(p14;q32) patient lies approximately 5 cM proximal to the 13q deletion region. Nevertheless, it overlaps the deleted segments of five patients showing interstitial deletions in the proximal 13q32 region and mild to moderate MR.13 Consistent with the published deletion cases, our patient displays only MR without severe malformation. Therefore, a gene responsible for mild MR may reside in the breakpoint region defined by YAC 737f10. Interestingly, three cDNA clones have been mapped to this YAC.15 Experiments are under way to see whether the breakpoint is located within or close to these coding sequences.
YAC 882b4 which spans the 13q21 breakpoint in a Moebius syndrome patient is of particular interest. Until now, two unrelated cases with MBS have been published where similar 13q regions were found to be rearranged. Ziter et al 16described a t(1;13)(p34;q13) cosegregating with MBS in seven members of a three generation family. Furthermore, a deletion of band 13q12.2 was found in a 2½ year old girl with MBS.17 Although these chromosome abnormalities seem to be located proximal to YAC 882b4, this does not rule out the possibility that the same gene is affected in all patients. Unfortunately, molecular mapping data are not available for the published cases. Upon FISH mapping a large number (>40) of DBCRs on different human chromosomes, we have observed that breakpoint localisation by banding analysis alone can be several bands out. Alternatively, a second gene causing Moebius syndrome might be located in the breakpoint region. Although we have mapped the breakpoint on 13q21 of the MBS patient, we cannot exclude that the aberrant phenotype is the result of the other autosomal breakpoints involved in this complex rearrangement. Molecular studies are in progress to study the MBS associated breakpoints of our patient in more detail, and breakpoint spanning clones are also being used to search for rearrangements in unrelated MBS patients with apparently normal karyotypes.
Several explanations may exist for the clustering of DBCRs in specific chromosome regions and for the occurrence of microdeletions in a breakpoint region. Genome mapping and sequencing projects have identified local duplications of (pseudo)genes and particular repetitive elements. It is well known that intrachromosomal recombination between nearby homologous sequences can result in deletion/duplication of intervening sequences. Accumulating evidence suggests that reciprocal microdeletion/duplication events are the molecular mechanism underlying several common human genetic disorders, including Williams-Beuren syndrome on 7q11.2, Smith-Magenis syndrome and Charcot-Marie-Tooth type 1A (CMT1A) on 17p11.2, Kallmann syndrome on Xp22.3, and colour blindness, haemophilia A, Hunter syndrome, and Emery-Dreifuss muscular dystrophy, all on Xq28.18-21
The human genome is estimated to contain more than one million repetitive sequences, ranging from the high copy number retroposon families Alu and L1 to rarely repeated or duplicated elements. In addition, there are many gene families and pseudogenes. Ectopic pairing of such repetitive elements may drive chromosome dynamics by bringing distant regions into recombinational contact. Because of the high repeat number in complex genomes, the chance of chromosome rearrangements caused by illegitimate recombination is relatively large even for regions with low recombination frequencies.22Recombination between homologous repeats is not necessarily limited to the same chromosomal locus. Regions which are highly enriched with Alu, L1, or other repeats may also serve as preferred sites of chromosomal translocations.
In conclusion, we have generated a molecular cytogenetic tool that makes it possible to characterise chromosomal abnormalities rapidly and in fine detail. Our YAC panel has added a new dimension to the MCN and will greatly facilitate the systematic clinical, cytogenetic, and molecular analysis of DBCRs. In particular, FISH mapping of breakpoint spanning clones in MR patients with de novo balanced chromosome rearrangements may pave the way for the positional cloning of new genes that are critical for brain development and function.
Acknowledgments
We wish to thank the German Resource Centre at the MPI for providing the YAC clones, and the diagnostic laboratories of the Mendelian Cytogenetics Network for providing us with lymphoblastoid cells. We also thank Kristine Kingsley, Susanne Freier, and Dietmar Vogt for excellent technical assistance and Birgit Schnieders for helpful discussion. This work was supported by the German Genome Programme/Deutsche Forschungsanstalt für Luft-und Raumfahrt eV (grant No 4763) and the EU-Commission (BMH4-CT97-2268).
References
Footnotes
-
↵* Present address: Department of Human Genetics, Leiden University, The Netherlands.