Article Text

Abstract
Transient neonatal diabetes mellitus (TNDM) is a rare condition which presents with intrauterine growth retardation, dehydration, and failure to thrive. The condition spontaneously resolves before 1 year of age but predisposes patients to type 2 diabetes later in life. We have previously shown that, in some cases, TNDM is associated with paternal uniparental disomy (UPD) of chromosome 6 and suggested that an imprinted gene responsible for TNDM lies within a region of chromosome 6q.
By analysing three families, two with duplications (family A and patient C) and one with several affected subjects with normal karyotypes (family B), we have further defined the TNDM critical region. In patient A, polymorphic microsatellite repeat analysis identified a duplicated region of chromosome 6, flanked by markers D6S472 and D6S311. This region was identified on the Sanger Centre’s chromosome 6 radiation hybrid map (http://www.sanger.ac.uk/HGP/Chr6) and spanned approximately 60 cR3000. Using markers within the region, 418 unique P1 derived artificial chromosomes (PACs) have been isolated and used to localise the distal breakpoints of the two duplications.
Linkage analysis of the familial case with a normal karyotype identified a recombination within the critical region. This recombination has been identified on the radiation hybrid map and defines the proximal end of the region of interest. We therefore propose that an imprinted gene for TNDM lies within an 18.72 cR3000 (∼5.4 Mb) interval on chromosome 6q24.1-q24.3 between markers D6S1699 and D6S1010.
- transient neonatal diabetes mellitus
- chromosome 6
- duplication
Statistics from Altmetric.com
Transient neonatal diabetes mellitus (TNDM) was first documented in 1852 by Kitselle who described the symptoms of his son.1 Patients present with intrauterine growth retardation and within the first few weeks of life show signs of dehydration, failure to thrive, and hyperglycaemia. Endogenous insulin levels are usually low or undetectable and most patients require exogenous insulin therapy for a median duration of three months. The incidence of the disorder is approximately 1 in 400 000 live births.2
The condition spontaneously resolves before 1 year of age, however some patients develop diabetes again later in life. Following the initial recovery, patients are clinically normal with normal blood glucose levels and are generally no longer monitored unless diabetes recurs. Hence it is difficult to comment on whether there is a continuing subclinical diabetes in the period between recovery and relapse. However, three patients who developed diabetes again in their teens have been investigated with the following results: all three were negative for islet cell antibodies; one showed insulin resistance following an oral glucose tolerance test; and one had periods of hyperglycaemia associated with infections before finally relapsing. Additionally, two younger patients who do not yet have diabetes also have periods of hyperglycaemia associated with infection and a third has normal glucose levels but raised C peptide levels indicative of insulin resistance.2 These findings, though not conclusive, suggest there may be subclinical features of diabetes following initial recovery in some TNDM patients.
In 1995, we reported two cases of TNDM associated with paternal uniparental disomy of chromosome 6 (UPD 6, patients D and E in table1).3 At that time there were two other documented cases of paternal UPD 6 (patients F and G), one of whom had hyperglycaemia but died two weeks after birth.4 The other showed no symptoms of TNDM.5 Since then, three more cases of TNDM with paternal UPD (patients H, I, and J) have been described6 7 (Gardner et al, in preparation) and one case of paternal UPD 6 was discovered fortuitously when the patient was 9 years old (patient K) with no history of TNDM.8 The explanation for the variable clinical picture with paternal UPD 6 is as yet unexplained, but as no specific neonatal tests for diabetes were performed on patient K and it is known that some patients do not require exogenous insulin therapy,9 hyperglycaemia may have been subclinical. The finding of paternal UPD 6 associated with TNDM gave the first indication that the causative gene(s) may be imprinted. In keeping with this hypothesis one case of maternal UPD 6 (patient L) has been reported with no symptoms of TNDM.10
Patients referred to in this study
We have previously described two families which provide further evidence of the involvement of an imprinted gene in TNDM.11 In family A, three subjects, the proband (patient A, table 1), her father, and her paternal grandmother, had a duplication of what was thought to be 6q22-q23 (now 6q24) but only the proband, who inherited the duplication from her father, had TNDM. Her father, who inherited the same duplication from his mother, showed no evidence of TNDM. FISH analysis using a whole chromosome 6 paint showed that the duplicated region was inserted into the short arm of chromosome 2. Family B included four subjects with TNDM in two generations (the proband is patient B in table 1). The disorder was inherited paternally in all cases and there was evidence of linkage to the marker D6S310, known to be duplicated in family A.11We are aware of two other subjects with paternal duplications involving chromosome 6q who also have TNDM (patients C and M)12 13 and a case of TNDM with a duplication of 6q of unknown origin (patient N).14 The further analysis of patient C is described below, whereas patients M and N were unavailable to us.
Using PCR amplification of polymorphic microsatellite repeats we partially characterised the extent of the duplication in family A, finding it to lie between markers D6S472 and D6S31111according to the Généthon map.15 In order to map the breakpoints more accurately and reduce the size of the critical region within which a gene for TNDM must lie, radiation hybrid mapped STS markers were used to isolate P1 derived artificial chromosomes (PACs) containing sequences from the region between D6S472 and D6S311. These PACs were ordered and used as FISH probes to analyse family A. These results, combined with the molecular mapping of a recombination in family B, allowed the TNDM critical region to be reduced and positioned at a more distal location.
Materials and methods
CA REPEAT ANALYSIS
DNA was extracted from whole blood using a salt precipitation method.16 Primers for polymorphic microsatellite repeat analysis were synthesised by Oswell DNA services or in house using an ABI 392 oligonucleotide synthesiser. Primer sequences were obtained from Généthon or GDB.15
Before amplification one of the primers was end labelled with32P-γATP using T4 polynucleotide kinase. Using the method of Hudson et al,17 DNA amplification was carried out in an ABI 9600 multiwell programmable thermal cycler. PCR products were separated by denaturing polyacrylamide gel electrophoresis and visualised by autoradiography.
ISOLATION OF PACS
PACs were isolated using the strategy described by Mungallet al,21 the only differences being that (1) 15 STSs were pooled and hybridised to the high density filters, and (2) the high density filters contained 16 × 384 well plates/filter in a 4 × 4 array. Primers used for generating labelled probes for hybridisation screening were produced by Genset.
FLUORESCENCE IN SITU HYBRIDISATION
Following semisynchronisation with Fdu and release with thymidine, chromosomes were prepared by standard techniques.18
PAC DNA was isolated using a rapid alkaline lysis method and the DNA labelled with digoxygenin using a nick translation kit (Amersham/Boehringer Mannheim). A modified protocol of the method of Pinkel et al 19 was used for FISH hybridisation and washing. Chromosomes were counterstained with DAPI and viewed through a Zeiss Axiophot microscope. Images were captured using a Photometrics 200 cooled CCD camera and enhanced using Extensions for QUIPS Smart Capture software package (Vysis). For each probe a minimum of five cells were captured and the results were corroborated by an independent observer.
Results
ISOLATION OF PAC CLONES
A 61.13 cR3000 region was mapped between markers stSG7277 at position 528.57 and stSG9891 at position 589.81 on the radiation hybrid map (table 2, Mungall et al, in preparation). The radiation hybrid map and marker data are contained within the 6ace database (http://www.sanger.ac.uk/HGP/Chr6). D6S472 was the most proximal marker used for (CA)n repeat analysis and D6S311 the most distal. They were located on the radiation hybrid map at positions 531.14 and 586.39 respectively. The boundary of the region to be mapped was then extended slightly (528.57-589.81, table 2) to ensure the whole region of interest was included and allow for ordering errors (the order of D6S472 and D6S1722 on the radiation hybrid map differs with the order according to the Généthon map15). A total of 165 markers within the region of interest were used to screen a PAC library made available to the Sanger Centre by Pieter de Jong.20 21 A total of 418 unique PAC clones were isolated from this region and have been assembled into contigs using hybridisation and fingerprinting data (http://www.sanger.ac.uk/HGP/Chr6).
Markers, radiation hybrid map positions, and positive PACs used in this study
IDENTIFICATION OF DISTAL BREAKPOINTS
Initially, four PAC clones (323P13, 333N19, 328C24, and 337A6)20 were selected as FISH probes. The first two hybridise to markers at the proximal and the second two to markers at the distal end of the critical region (table 2). When these four PACs were hybridised to metaphase chromosomes from a member of family A, the proximal clones 323P13 and 333N19 hybridised to both chromosomes 6 and to the insertional duplication on chromosome 2. The distal clone 328C24 only hybridised to the two normal chromosomes 6. This indicates that although the distal breakpoint is within the targeted region, the duplication extends further proximally than the region covered by the chosen PACs. (Clone 337A6 hybridised to chromosome 20q11.2 as well as chromosome 6.) Additional PAC clones were used to narrow the region containing the distal breakpoint. This was found to lie between clone 430G4 which hybridised to marker stSG10998 at position 569.62 and clone 20H10 which hybridised to stCHLC.GATA41E03, stSG12407, and stSG12423 at position 571.22 on the radiation hybrid map (table 2). The distal breakpoint of a second duplication, in patient C,13 was found to lie between clones 448H8 and 28C20 which hybridised to markers stSG12407 and stSG12423 at position 571.22 and stSG11104 and stSG11157 at position 571.22, respectively. This did not refine the area of interest. While the two breakpoints are not in exactly the same place, they are in close proximity. It may be that this region of the genome is prone to breakage and it would be interesting to determine the exact position of any other breakpoints within this region to see whether there is a breakpoint cluster.
IDENTIFICATION OF THE PROXIMAL LIMIT OF THE CRITICAL REGION
Previously, linkage has been shown between TNDM and the polymorphic marker D6S310 in family B.11 We now have DNA samples from 14 members of this family and haplotype analysis using polymorphic markers from the region between D6S472 and D6S311 has allowed us to identify a TNDM associated haplotype.
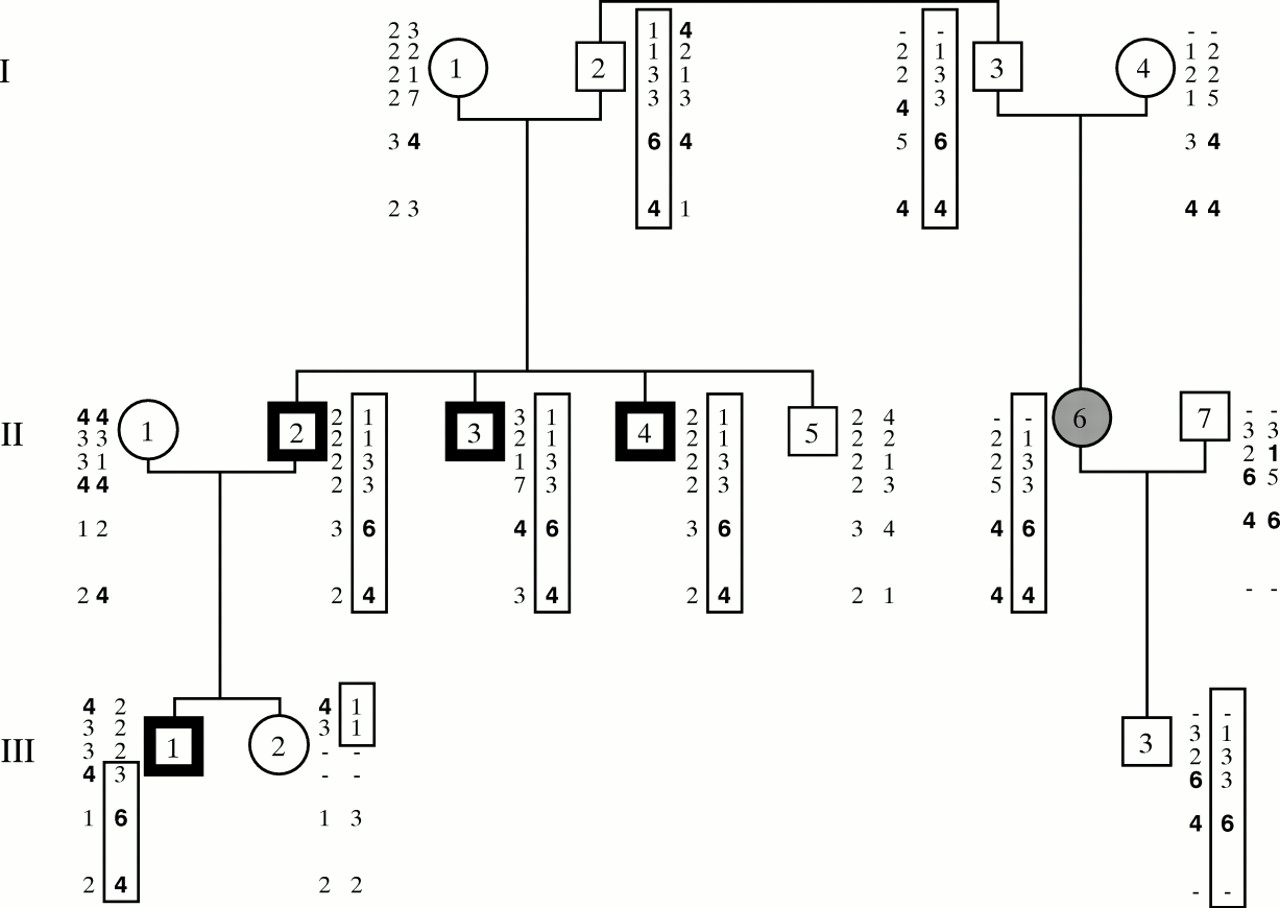
Fig 1 shows that the same paternal haplotype has been inherited by the three males with TNDM in generation II (II.2, II.3, and II.4) while the unaffected male (II.5) has inherited a different paternal haplotype. Their female cousin (II.6) also inherited the high risk haplotype from her father. Although she was not diagnosed as having TNDM, she is older than her affected cousins and was born before the family was alerted to TNDM. She is reported to have had a birth weight of only 2300 g at term and a “very difficult” neonatal period. This could have been because of TNDM, which spontaneously resolved without the need for exogenous insulin therapy. She developed gestational diabetes and now has type 2 diabetes which is treated by diet alone. Her son (III.3) did not have TNDM, but has the at risk haplotype. However, unlike his affected cousin and uncles, he inherited it from his mother.

{kind=link}
The pedigree of family B showing the haplotypes determined by PCR amplification of polymorphic microsatellite repeats. The six sets of results are, from top to bottom, D6S472, D6S975, D6S1699, D6S314, D6S310, D6S1703. The boxed haplotype is the high risk haplotype, all or part of which has been inherited, from their father, by all subjects thought to have had TNDM. III.3 inherited the high risk haplotype from his mother and as expected did not have TNDM. III.2 only inherited part of the high risk haplotype but does not have TNDM, indicating that the gene lies distal to her recombination. The diagnosis that III.2 did not have TNDM is, we believe, an accurate one as the family and clinicians involved were all aware of the condition following the diagnosis of her father, uncles, and older brother. II.6 is shaded in grey as she is thought to have had undiagnosed TNDM (see text).
The haplotypes of III.1 and III.2 indicate that recombinations have occurred. As III.2 does not have TNDM and her brother (III.1) does, the gene must lie within a region of the paternal haplotype not shared by these sibs. This places the gene distal to III.2’s recombination, that is, distal to the marker D6S975 on the Généthon map.15However, III.1 had TNDM and only has the high risk haplotype for the region distal to D6S1699. Therefore, the putative TNDM gene must lie distal to D6S1699 but within the region duplicated in family A. Thus, D6S1699 defines the proximal limit of the critical region as position 552.50 on the radiation hybrid map and considerably reduces the size of the critical region.
CHROMOSOMAL LOCALISATION OF THE CRITICAL REGION
To assign the critical region to a chromosomal location the clones 430G4 and 337E9 were hybridised to high resolution chromosomes from a person with a normal karyotype. 430G4 is the most distally duplicated clone (contains marker stSG10998 at position 569.62 on the radiation hybrid map) and 337E9 (contains D6S1699) marks the proximal boundary at position 552.5 on the radiation hybrid map. 337E9 hybridised to band 6q24.1 while 430G4 hybridised to band 6q24.3. This indicates that the critical region is more distal than originally suggested by microsatellite mapping and karyotype analysis of the insertional duplication in family A.11
Discussion
The first indication that a gene for TNDM may be located on chromosome 6 came when patient E was found to have a ring chromosome 6 of maternal origin and two normal chromosome 6 homologues, both of paternal origin. The patient also had TNDM. This finding prompted the analysis of two more TNDM patients, one of whom also had paternal UPD 6, leading to the hypothesis that an imprinted gene for TNDM is located on chromosome 6.3 Since then the region of interest has been sequentially reduced from the whole of chromosome 6 to a small 18.72 cR3000 region within 6q24.
Parameters for the human genome have been established by Morton,22 which enable centiray units which are dose dependent to be expressed in terms of megabases. As the total length of the radiation hybrid map of chromosome 6 is 636.21 cR3000, and the physical map length is 183 Mb,22 there are approximately 3.48 cR3000/Mb. The region of 61.13 cR3000 mapped initially is approximately 17.6 Mb while the critical region of 18.72 cR3000 is approximately 5.4 Mb. Although radiation breakage hotspots can result in a non-linear relationship between the radiation hybrid map and physical distance, our figure of 5.4 Mb is consistent with the Genetic Location Database estimate of physical distance between D6S1010 and D6S1699 of ∼6.4 Mb.23
FISH analysis using PACs from the TNDM critical region and high resolution chromosome preparations has refined the location of the TNDM gene to 6q24.1-q24.3. It is known to be difficult to map small chromosomal insertions accurately owing to the three band uncertainty principle described by Savage,24 and originally karyotype analysis together with the map location of microsatellite markers in family A suggested the critical region lay within G dark 6q22 and G light 6q23. The new results show the insertional duplication in family A comprises G light 6q23 and G dark 6q24 rather than 6q22. Similarly, the duplication in patient C, originally thought to involve 6q16-q21 on karyotype analysis and reassigned to 6q22-q23 upon microsatellite repeat analysis,13 is now known to extend into 6q24 after analysis with FISH probes from the TNDM critical region. These cases serve to illustrate the greater accuracy over other techniques of FISH mapping with computer enhanced DAPI banding and may explain the apparent discrepancy between the 6q24 location of the TNDM gene and the published 6q21-q23 location of the duplication in patient N,14 who is not yet available for FISH analysis.
The phenotype of patients A and C who both have paternal duplications is mild considering that the duplications are both cytogenetically visible and therefore relatively large. Patient C has TNDM, macroglossia, and mild dysmorphic features,13 whereas patient A has TNDM, macroglossia, and mental retardation, probably resulting from prolonged severe dehydration as a result of untreated TNDM in the neonatal period.11 The phenotype of patient N who has a duplication of unknown parental origin is more complex with hyperglycaemia being just one feature of many.14 A fourth case (patient M) of neonatal hyperglycaemia was described as part of a complex phenotype resulting from partial paternal trisomy of 6q23-qter.12
Two additional cases of partial paternal trisomy of 6q including the newly defined critical region have been reported. Patient O was trisomic for 6q23-qter but died at birth.25 Patient P has a duplication of 6q24-qter resulting in congenital anomalies but with no mention of neonatal diabetes.26 Analysis of material from this case using the PAC clones may further localise the proximal breakpoint, but so far material from this patient has been unavailable for study.
A considerable resource of clones from the TNDM critical region has now been established. Gap closure methods will be used in order to produce sequence ready contigs as part of the chromosome 6 mapping and sequencing project. This will also aid the mapping of the TNDM gene, as a complete contig of the region will soon be available.
We hypothesise that overexpression of an imprinted gene results in TNDM and this hypothesis is supported by the finding of paternal UPD 6 or large paternal duplications of 6q in some TNDM patients. Hence it is likely that other TNDM patients will have duplications involving 6q24. We are currently using PACs which hybridise to markers within the newly defined critical region as FISH probes to analyse TNDM patients in whom a structural abnormality of chromosome 6 has not been identified by karyotype and microsatellite repeat analysis. We hope that this will result in the detection of smaller structural anomalies and will further refine the region containing the gene. Once a critical region of a size suitable for gene hunting techniques has been characterised it will be possible to isolate candidate genes for further study.
Several known genes have been localised to a region of 6q including band 6q24 according to the Genome Database, though none stand out as obvious candidate genes. However, several previously or potentially implicated genes do map to chromosome 6 but not to the correct cytogenetic location: IDDM15 (insulin dependent diabetes mellitus 15, 6q21),27 IDDM5 (insulin dependent diabetes mellitus 5, 6q25),28 IDDM8 (insulin dependent diabetes mellitus 8, 6q27),29 IGF2R (insulin-like growth factor 2 receptor, 6q25.3),30 and PDNP1 (PC-1 phosphodiesterase I, 6q22-q23).31
In conclusion we have generated a resource of PACs from a 61.13 cR3000 region of 6q and have defined an approximately 5.4 Mb (18.72 cR3000) region of chromosome 6q24.1-q24.3 in which the gene for TNDM must lie.
Acknowledgments
We would like to thank Sally Ranby and Carol Edwards for assistance with the library screening; Sean Humphray and Rose Heathcott for the fingerprinting analysis; members of the resource laboratory at the Sanger Centre for the provision and maintenance of the library and filter grids; Sarah Beal and Christine Joyce for their help with the FISH analysis; Drs M Kibirige, A Ferguson, and J Zlotogora for referring families A, B, and C respectively and Dr Andy Collins for help with estimating the size of the critical region. RJG is supported by grants from the British Diabetic Association and the Wessex Medical Trust while the work at the Sanger Centre is supported by the Wellcome Trust.